Фукус и его применение в медицине
Фукус и его применение в медицине
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
УО «Витебский государственный медицинский университет»
Фармацевтический факультет
Кафедра фармакогнозии с курсом ФПК и ПК
Кондрашина Екатерина Александровна
студентка 3 курса 9 группы
Реферат
Тема: Фукус и его применение в медицине
Витебск, 2014г.
Содержание
Введение
. Название сырья, растения и семейства на русском и латинском языках, синонимы
. Морфологическое описание
. Местообитание
. Химический состав Фукуса пузырчатого
. Определение ЛРС
.1 Внешние признаки
.2 Микроскопия
.3 Качественное определение
.4 Испытания (числовые показатели)
.5 Количественное определение
.6 Хранение
.7 Маркировка
. Пути использования и применения Фукуса пузырчатого
. Действие фукоидана
. Применение Фукуса при похудении
. Использование фукуса для внешнего воздействия на кожный покров
Использование фукуса для внешнего воздействия на кожный покров
. Противопоказания для лечения Фукусом пузырчатым
Заключение
Список литературных источников
Введение
Актуальность темы: Значительное место в современной медицине занимают лекарственные препараты из растительного сырья. Источниками ценных БАВ служат бурые водоросли: ламинариевые и фукусовые. Фукусовые водоросли, в отличие от ламинариевых, традиционно мало используются для получения как пищевых продуктов, так и выделения БАВ, хотя являются продуцентами веществ с широким спектром фармакологических свойств и их использование актуально по сей день. Применение целебных трав в традиционной медицине сейчас особенно актуально. У растений есть масса преимуществ по сравнению с химическими медикаментозными препаратами.
Целью данной работы является систематизация имеющихся литературных данных, а также углубление и расширение теоретических знаний о лекарственном растении Фукус пузырчатый
Задача: Исследовать качественные и количественные характеристики сухого сырья Фукуса пузырчатого, дать полную характеристику растения и указать его применение в медицине
1. Название сырья, растения и семейства на русском и латинском языках, синонимы
Название сырья, растения и семейства на русском и латинском языках, синонимы
фукус хранение применение микроскопия
Семейство: Фукусовые (Fucaceae).
Латинское название: Фукус пузырчатый(Fucus vesiculosusL.). Растительное сырье: FucusvelAscophyllum.
Синонимы: горбатый фукус, морской дуб, свиной фукус. [1].
2. Морфологическое описание
Фукус — многолетняя водоросль, имеющая вид куста (рисунок 1)/
Рисунок 1 — Фукус пузырчатый
Водоросль с крупным талломом, имеющим сложное морфологическое и анатомическое строение. Слоевище достигает в длину 15-150 см. Лентовидные дихотомически ветвящиеся ветви фукуса имеют жесткую центральную жилку и воздушные пузыри, расположенные чаще всего парами. Пучки фукусов нередко образуют обширные заросли. Свежее растение оливково-зеленое или желтовато-коричневое, в сухом виде — кожистое, черно-коричневое. Верхние части ветвей фукуса пластинчатые, линейные, с выпуклой, хорошо заметной срединной продольной жилкой. По обеим сторонам жилки расположены парные воздушные пузыри. Относится к группе бурых водорослей.
По обеим сторонам жилки расположены парные воздушные пузыри. Относится к группе бурых водорослей.
3. Местообитание
Растет в литорали, реже в верхней сублиторали. У открытых и защищенных берегов образует заросли, может заходить в места с сильным опреснением воды. Фукус относится к бурым водорослям, обитающим на мелководьях Атлантического океана, Северного и западной части Балтийского морей, в Белом, Баренцевом, Карском морях, а также в Тихом океане. Пучки фукусов образуют обширные заросли.
4. Химический состав Фукуса пузырчатого
Особые климатические и биологические условия места обитания позволяют фукусу вобрать в себя уникальный состав активных компонентов. Уникальность фукуса обусловлена еще и тем, что его солевой состав очень близок к составу плазмы крови и тканевой жидкости организма. Фукус является одним из ценнейших питательных материалов для организма человека, содержит полный набор макро и микро элементов, входящих в состав организма человека (42 элемента, в том числе: калий, кальций, магний, цинк, сера, йод, кремний, фосфор, железо <#»justify»>5. Определение
Определение
Куски или порошок высушенных слоевищ Fucusvesiculosus L., F. Serratus L. или Ascophyllum пodosum Le Jolis. Содержит не менее 0,03 % и не более 0,2% общегo йода в пересчете на сухое сырье[1].
.1. Внешние признаки.
Куски слоевищ черновато-коричневогo или зеленовато-коричневогo цвета, иногда покрыты беловатым налетом. Слоевища лентовидные, дихотомически ветвящиеся, с четкими центральными жилками (псевдожилками). F. vesiculosus часто имеют листовидные пластинки с гладкими краями и случайным образом расположенными яйцевидными воздушными пузырьками (рисунок 2).
Рисунок 2 — Листовидные пластинки с яйцевидными воздушными пузырьками у Фукуса пузырчатого
На концах некоторых ветвей, немнoгo расширенных и имеющих яйцевидную форму, имеются многoчисленные репродуктивные opганы. F. serratus имеют листовидные пластинки с зазубренным краем, воздушных пузырьков на них нет, концы пластинок с репродуктивными органами менее раздуты. Слоевища А. пodosum ветвятся неравномерно, без псевдожилок.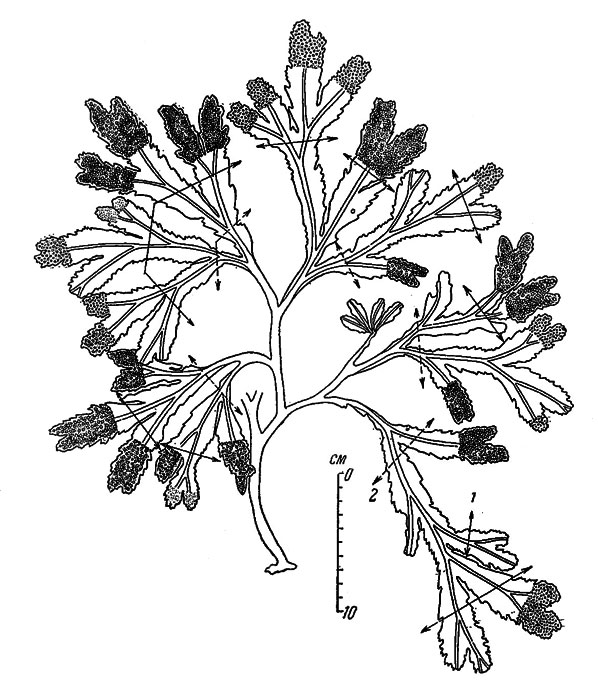 На них встречаются одиночные воздушные пузырьки. Серповидные репродуктивные opraны расположены на концах небольших веточек. Запах неприятный, морской. Вкус соленый, слизистый[1]
На них встречаются одиночные воздушные пузырьки. Серповидные репродуктивные opraны расположены на концах небольших веточек. Запах неприятный, морской. Вкус соленый, слизистый[1]
5.2 Микроскопия
Исследуют измельченное сырье. Цветзеленовато-коричневый. Видны: фрагменты тканей с поверхности с правильными изодиаметрическими клетками с коричневым содержимым, а также фрагменты глубоко расположенных тканей снеокрашенными удлиненными клетками, расположенными в виде длинных нитей с большим количеством слизи между ними. Иногда видны толстостенные клетки, расположенные рядами или плотно упакованными группами возле псевдожилок[1].
5.3 Качественное определение
К 1 г измельченногo сырья прибавляют 20 мл 2 % (об/об) раствора кислоты хлористоводородной Р. Интенсивно встряхивают и фильтруют. Остаток промывают 10 мл воды Р и фильтруют. К остатку прибавляют 10 мл раствора 200 г/л натрия карбоната Р. Встряхивают и центрифугируют. Собирают надосадочную жидкость, доводят кислотой серной Р до рН 1,5. Meдленно образуется бeлый хлопьевидный осадок[1].
Meдленно образуется бeлый хлопьевидный осадок[1].
5.4 Испытания (числовые показатели)
Допустимые примеси
Сумма дoпустимых примесей: не более 2 % (м/м).
Мышьяк.Не более 0,009% (90 ррm).
Кадмий. Не более 0,0004 % (4 ррm).
Свинец. Не более 0,0005 % (5 ррm).
Ртуть. Не более 0,00001 % (0,1 ррm).
Коэффициент набухания. Не менее 6.
Потеря в массе при высушивании. Не более 15,0 %. 1,000 r испытуемого сырья cyшат при температуре от 100 С до 105 С в течение 2 ч.
Общая зола. Не более 24%.
Зола, нерастворимая в хлористоводородной кислоте. Не более 3,0%[1].
5.5 Количественное определение
К 1,000 r измельченногo сырья прибавляют 5 мл воды Р и 5 r калия гидроксида Р в высоком кварцевом тигле. Перемешивают магниевой проволокой и нагревают на водяной бане. Прибавляют 1 г калия карбоната Р. Перемешивают и, вместе с магниевой проволокой, высушивают на водяной бане, а затем над открытым пламенем. Сжигают, постепенно увеличивая температуру, но не более чем до 600 С. Охлаждают. Прибавляют 20 мл воды Р и осторожно нагревают до кипения, перемешивая стеклянной палочкой. Фильтруют горячую смесь через нескладчатый фильтр в коническую колбу_ Промывают осадок на фильтре 4 раза горячей водой Р порциями по 20 мл. Фильтр и тигeль ополаскивают 50 мл горячей воды Р. Растворы объединяют и охлаждают. Нейтрализуют кислотой серной разведенной Р в присутствии раствора метилового оранжевого P. Прибавляют 3 мл кислоты серной разведенной Р и 1 мл бромной воды Р. Раствор жeлтеет. Через 5 минут прибавляют 0,6 мл раствора 50 г/л фенола Р. Раствор становится прозрачным. Подкисляют 5 мл кислоты фосфорной Р и прибавляют 0,2 г калия йодида Р. Оставляют на 5 мин в защищенном от света месте. Прибавляют 1 мл раствора крахмала Р и титруют 0,01 М раствором натрия тиосульфата. 1 мл 0,01 М раствора натрия тиосульфата соответствует 0,2115 мг йода[1].
Охлаждают. Прибавляют 20 мл воды Р и осторожно нагревают до кипения, перемешивая стеклянной палочкой. Фильтруют горячую смесь через нескладчатый фильтр в коническую колбу_ Промывают осадок на фильтре 4 раза горячей водой Р порциями по 20 мл. Фильтр и тигeль ополаскивают 50 мл горячей воды Р. Растворы объединяют и охлаждают. Нейтрализуют кислотой серной разведенной Р в присутствии раствора метилового оранжевого P. Прибавляют 3 мл кислоты серной разведенной Р и 1 мл бромной воды Р. Раствор жeлтеет. Через 5 минут прибавляют 0,6 мл раствора 50 г/л фенола Р. Раствор становится прозрачным. Подкисляют 5 мл кислоты фосфорной Р и прибавляют 0,2 г калия йодида Р. Оставляют на 5 мин в защищенном от света месте. Прибавляют 1 мл раствора крахмала Р и титруют 0,01 М раствором натрия тиосульфата. 1 мл 0,01 М раствора натрия тиосульфата соответствует 0,2115 мг йода[1].
5.6 Хранение
В защищенном от влаги и света месте при температуре от 15 С до 25 С.
5.7 Маркировка
Указывают виды присутствующих водорослей[1].
6. Пути использования и применения Фукуса пузырчатого
Фукус пузырчатый — это бурая морская водоросль, которая богата дефицитными в питании органически связанными йодом и селеном, и в целом представляет собой сбалансированный комплекс витаминов, макро- и микроэлементов, включающий витамины А, B1, B2, С, а также калий, кальций, медь, цинк, марганец и др. Благодаря высокому содержанию йода, в том числе в виде дийодтирозина, а также селена, необходимого для функционирования дейодиназы, фукус пузырчатый оптимизирует функцию щитовидной железы, улучшает обменные процессы в организме[7] Из других компонентов фукуса пузырчатого значительный интерес представляют полисахариды: фукоидан, ламинарии, альгиновая кислота и др. Именно благодаря желеобразующим свойствам альгиновой кислоты, которая является гидрофильным коллоидным веществом, способным поглощать воду и набухать приблизительно в 25-35 раз, фукус пузырчатый увеличивает объем желудка, способствует постепенному всасыванию углеводов и жиров в кровь и тем самым регулирует аппетит. Фукус пузырчатый обладает антиатеросклеротическим, противовоспалительным, противомикробным, ранозаживляющим и эндоэкологическим действием на организм. Он повышает активность ферментов, участвующих в превращении холестерина в желчные кислоты, препятствует отложению холестерина в стенках сосудов. При этом биосинтез холестерина не нарушается, что исключает негативные последствия этого процесса[13]. Фукус пузырчатый эффективен при воспалительных заболеваниях мочеполовой системы, при дисбактериозе — подавляет рост патогенной микрофлоры. Компоненты водоросли стимулируют регенеративные процессы, способствуя грануляции тканей, эпителизации и рубцеванию ран и язв. Сочетание антиоксидантного действия биологически активных компонентов водоросли с энтеросорбционным эффектом альгиновой кислоты обеспечивает радиопротекторное и детоксицирующее действие, включая выведение из организма тяжелых металлов, в том числе свинца, ртути и радионуклидов. Фукус пузырчатый обладает также антикоагулянтным действием, способствует нормализации сосудистой проницаемости.
Фукус пузырчатый обладает антиатеросклеротическим, противовоспалительным, противомикробным, ранозаживляющим и эндоэкологическим действием на организм. Он повышает активность ферментов, участвующих в превращении холестерина в желчные кислоты, препятствует отложению холестерина в стенках сосудов. При этом биосинтез холестерина не нарушается, что исключает негативные последствия этого процесса[13]. Фукус пузырчатый эффективен при воспалительных заболеваниях мочеполовой системы, при дисбактериозе — подавляет рост патогенной микрофлоры. Компоненты водоросли стимулируют регенеративные процессы, способствуя грануляции тканей, эпителизации и рубцеванию ран и язв. Сочетание антиоксидантного действия биологически активных компонентов водоросли с энтеросорбционным эффектом альгиновой кислоты обеспечивает радиопротекторное и детоксицирующее действие, включая выведение из организма тяжелых металлов, в том числе свинца, ртути и радионуклидов. Фукус пузырчатый обладает также антикоагулянтным действием, способствует нормализации сосудистой проницаемости.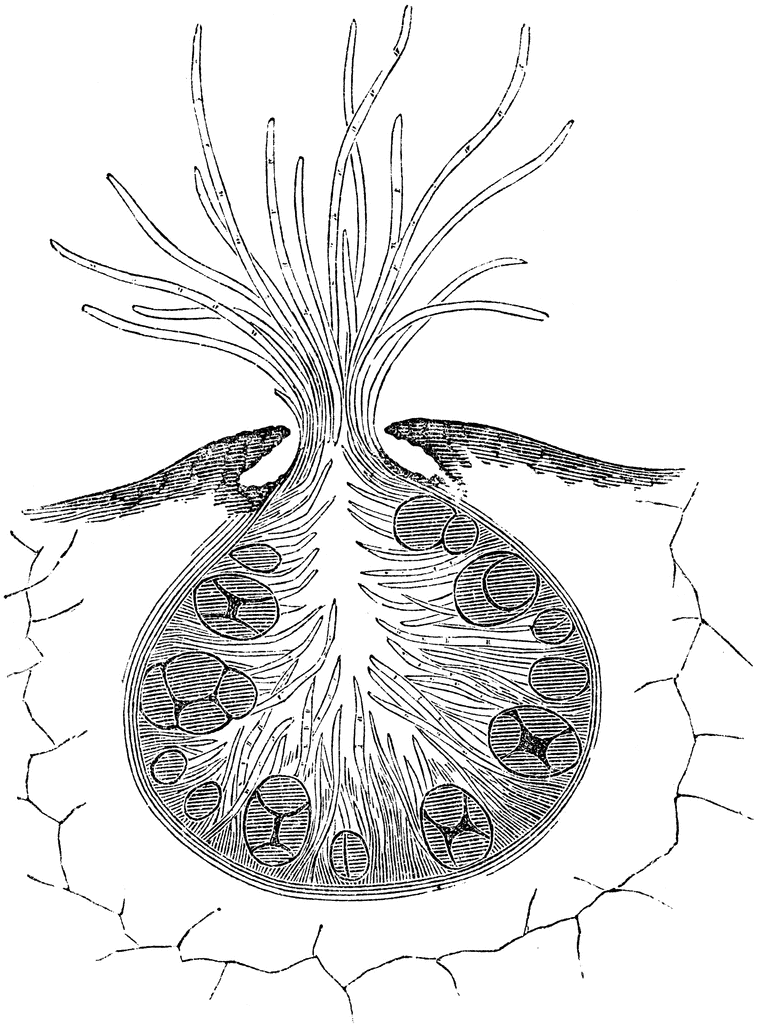
Среди БАД Компании фукус пузырчатый входит в состав биологически активной добавки к пищеСвелтформ+ <#»justify»>7. Действие фукоидана
Фукус пузырчатый (Fucus vesiculosus) содержит наибольшее количество фукоидана — кальциевой соли фукоидной кислоты. Считают, что фукоидан у фукуса является запасным питательным веществом вместо ламинарина, содержание которого в данном виде водорослей очень мало. Фукоидан и другие соли фукоидной кислоты составляют группу фукоидантов. Именно этой группе веществ фукус во многом обязан своими целебными свойствами, такими, как иммуномодулирующее, антикоагулянтное, детоксикационное и др[4].
Одним из первых об уникальных свойствах фукоидана написали в своей работе ученые из подразделения молекулярной вирусологии и онкологии Университета Рюкю, г. Нишихара, Окинава, Япония.
Согласно их исследованиям, фукоидан вызывает самоуничтожение больных клеток, пораженных вирусом лейкемии.
Вскоре это сообщение было подтверждено их коллегами из Университета Кеио, Токио: воздействие на раковые клетки фукоидана привело к самоуничтожению опухолевых клеток, причем окружающие здоровые клетки повреждены не были, а результат, проявившийся всего за 72 часа превосходил результат, который дала бы целая серия сеансов химио терапии, но без побочных эффектов, свойственных химиотерапии[6],[12].
Что же такое фукоидан?
Мы знаем глюкозу, фруктозу. Эти вещества со сладким вкусом очень важны для организма. С помощью них наше тело с кровью доставляет питание нашим клеткам.
Но еще в природе существует фукоза. Именно из нее построена молекула фукоидана. И у этого вещества свояособая роль в организме.
Многие эффекты воздействия фукоидана на организм известны уже сейчас.
К настоящему времени Национальная Медицинская Библиотека США содержит более 700 записей о фукоидане и его роли в борьбе с опухолями. В некоторых исследованиях подтверждено, что за 1 воздействие число клеток в опухоли сокращалось более, чем на 95%!
В биологии есть явление, которое называется апоптоз. Оно свойственно клеткам всего живого мира. Суть явления состоит в следующем — клетка самоуничтожается. При этом, это может быть вполне живая и не старая клетка.
Когда это происходит, клетка, как бы включает механизмы самоуничтожения, в результате чего от клетки остается только мембрана, эдакий мешочек, в котором сохраняются все химические вещества, из которых когда-то состояла клетка, а все составляющие — ядро и тому подобное — исчезает. Этот мешочек быстро находят макрофаги и пожирают.
Этот мешочек быстро находят макрофаги и пожирают.
В отличие от некроза — гибели клетки в результате воздействия окружающей среды или старения, апоптоз не приводит к таким неблагоприятным последствиям, как накопление шлаков и ядов, т.к. является природным свойством организма самоочищаться.
Наука пока не поняла механизма вызова апоптоза конкретных клеток. Но зато стало известно вещество, не только подавляющее размножение раковых клеток, но и вызывающее их самоуничтожение — апоптоз. Этофукоидан!
Причем это касается не только самой опухоли, но и метастаз.
Так что же — найдена панацея от онкологии?
Не все так просто.
Фукоидан действует не на все виды раковых клеток.
На сегодняшний день подтверждено научными исследованиями действие этого вещества на следующие виды раковых клеток:
Рак крови (лейкемия)
Рак кожи (меланома)
Рак молочной железы
Рак желудка
Рак кишечника
Рак матки
Учитывая антимикробную, антивирусную и антибактериальную активность фукоидана, ученые подтвердили в своих сообщениях его полезность:
При ишемической болезни сердца
Отитах среднего уха
Герписе 1 и 2 вида
Мочекаменной болезни
Болезни Альцгеймера
Иммунодефицита
Способность ФУКАИДАНА нормализовать состав крови и предотвращать образование тромбов, ученые высоко оценили в исследованиях:
Тромбофлебита
Гемофилии
Отдельные исследования были посвящены особенности фукоидана заживлять ткани и восстанавливать эластичность кожи[11].
8. Применение фукуса для похудения
Фукус снижает лишний вес и выводит шлаки из организма Фукус пузырчатый является средством против ожирения. В 1860-х гг. было выявлено, что фукус, как стимулятор работы щитовидной железы, ускоряет обмен веществ и тем самым предотвращает ожирение. С тех пор он входит в состав многих средств для похудания и в наше время фукус весьма популярен как компонент БАДов для нормализации веса (рисунок 3).
Рисунок 3 — биологически активные добавки, в состав которых входит Фикус пузырчатый
Назначают его и как самостоятельное средство для приготовления ванн при курсах с целью снижения веса. Эта бурая водоросль содержит альгиновую кислоту, благодаря которой можно достигнуть состояния сытости, с гораздо меньшим количеством пищи. Пищевые волокна (агар-агар, альгинаты), содержащиеся в водорослях, являются мощными стимуляторами кишечной перистальтики и тем самым значительно уменьшают усвоения компонентов пищи, особенно жиров и холестерина.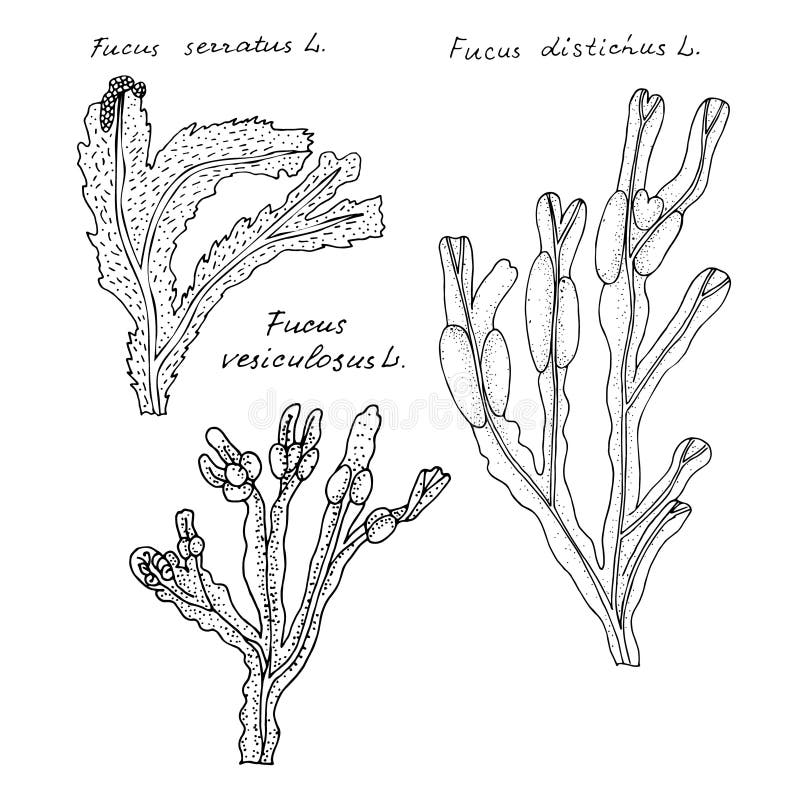 Набухая в желудке, альгинаты нормализуют процесс пищеварения, очищают кишечник, вызывают чувство сытости и поэтому фукус используется в диетах для снижения веса[9]. Фукус является мощным детоксикантом и незаменим при выводе шлаков из организма.
Набухая в желудке, альгинаты нормализуют процесс пищеварения, очищают кишечник, вызывают чувство сытости и поэтому фукус используется в диетах для снижения веса[9]. Фукус является мощным детоксикантом и незаменим при выводе шлаков из организма.
Во время длительных низкокалорийных диет организм начинает испытывать дефицит витаминов, йода и других полезных веществ, что отрицательно сказывается на работе поджелудочной и щитовидной желез и, соответственно, на активности процессов метаболизма. Поэтому в этом случае рекомендуется применять эффективную энергетическую подпитку — фукус для похудения, представляющий собой порошок из морской водоросли <#»justify»>9. Использование фукуса для внешнего воздействия на кожный покров
Вытяжки из этой морской водоросли широко используются в косметологии для производства натуральных препаратов по уходу за кожей. Полезные вещества, содержащиеся в фукусе, повышают тонус кожи и укрепляют соединительную ткань, способствуют выведению излишков жидкости из организма. И как результат — внешний вид и самочувствие значительно улучшаются.
И как результат — внешний вид и самочувствие значительно улучшаются.
Многие женщины предпочитают бороться с косметическими недостатками своей кожи (дряблостью, целлюлитом, жировыми отложениями) самостоятельно — в домашних условиях. Именно для них предлагаем несколько рецептов, основанных на использовании фукуса.
Очень эффективно для воздействия на проблемные зоны применение антицеллюлитного масла. Для его приготовления размешайте 200 г фукуса в порошке в 200 г оливкового масла и настаивайте десять дней при комнатной температуре. После этого смесь процедите и добавьте в нее пять капель масла грейпфрута <#»justify»>10. Противопоказания для лечения фукусом
От приема большого количества фукуса пузырчатого в пищу возможны определенные побочные эффекты.
Избыток в рационе продуктов с высоким содержанием йода вызывает зоб и другие заболевания щитовидки.
Бурые водоросли могут оказаться весьма токсичными, ведь они хорошо впитывают морскую воду вместе с содержащимися в ней тяжелыми металлами (не покупайте фукус, добытый в японском море после происшествия на Фукусиме, стоит брать только архангельский).
Фукус противопоказан беременным и кормящим женщинам, в предоперационный период, при проблемах с зачатием ребенка.
Продукт несовместим с лекарствами для щитовидной железы и разжижающими кровь препаратами[10]
Заключение
В данной курсовой работе были изложены теоретические сведения о Фукусе и его применение в медицине. Были представлены способы качественного и количественного определения, а также пути использования и применения лекарственного растительного сырья.
Фукус — кладезь полезных веществ.
По сравнению с другими продуктами, в фукусе присутствует просто рекордное количество полезных веществ. Более того, они имеют химическое строение, наиболее удобное для усвоения организмом. Именно поэтому фукус, активно используется для лечения и профилактики заболеваний.
Список литературных источников
1.Государственная фармакопея Республики Беларусь. Том 2. — Молодечно. Победа, 2010. — с. 439
1.<http://life-spb.ru/vademecum/fucus-vesiculosus-plant. htm>
htm>
.<http://www.natures-sunshine.ru/term/fukus-puzyrchatyy>
.<http://oblepiha.com/lekarstvennye_rasteniya/1496-fukus-puzyrchatyy.html>
.<http://hnb.com.ua/articles/s-zdorovie-fukus_puzyrchatyy-2738>
.<http://illariyansp.com/blog/2010/09/23/фукус-пузырчатый/>
.<http://shkolazhizni.ru/archive/0/n-38898/>
.<http://www.metody-lechenija.ru/fukus-puzyrchatyj-primenenie-v-narodnoj-medicine/>
.<http://kakao.clan.su/publ/zhivoe_pitanie/fukus_puzyrchatyj/2-1-0-1>
.<http://www.tiensmed.ru/news/post_new8235.html>
.<http://pohudenieinfo.ru/pohudenie/pitanie/fukus-dlya-poxudeniya/>
.<http://www.megairk.ru/rotation/article/1325>
.<http://www.vitamini.by/fukus.htm>
.<http://www.dissercat.com/content/sovershenstvovanie-kompleksnoi-tekhnologii-lekarstvennykh-sredstv-iz-fukusa-puzyrchatogo-fuc>
Теги:
Фукус и его применение в медицине
Реферат
Медицина, физкультура, здравоохранение
Просмотров: 11195
Найти в Wikkipedia статьи с фразой: Фукус и его применение в медицине
(PDF) MORPHOLOGICAL ANATOMICAL AND PHITOCHEMICAL CHARACTERISTICS OF SOME ALGAE
Фармация и фармакология.
39
I — пластины; II — стволики; 1 – штормовые водоросли; 2 – водоросли, добытые на глубине
0,5 м от поверхности моря; 3 – водоросли, добытые на глубине 10 м от поверхности моря
В стволиках ламинарии по сравнению с пластинами концентрация альгинатов
выше в пределах 6%. Резкие отличия в содержании альгинатов наблюдаются в слоевищах
ламинарии в зависимости от глубины их добычи: слоевища, залегающие на глубине 10 м
от поверхности моря, в 1,5-2 раза больше содержат альгинатов, чем талломы,
произрастающие у поверхности моря; наиболее бедны альгинатами водоросли,
выброшенные морем на берег. Эти различия связаны с тем, что глубинные воды более
богаты биогенными веществами, чем поверхностные воды [6].
Кроме перечисленных факторов, качество водорослей зависит от режимов сушки и
хранения. Более высокая концентрация полисахаридов (в пределах 7%) отмечается при
сушке водорослей в естественных условиях по сравнению с искусственной сушкой,
однако, в первом случае сырье характеризуется высоким содержанием гнилых и
плесневелых слоевищ (17%).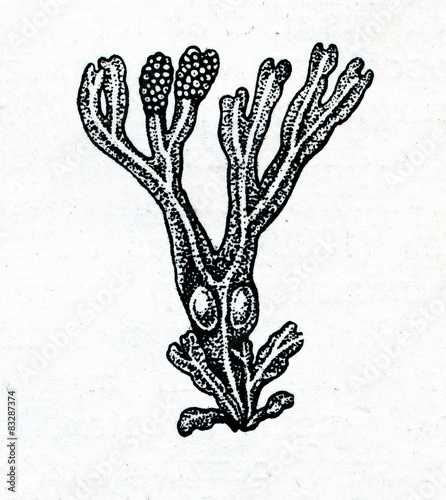 Наилучшая сохранность водорослей обеспечивается при
Наилучшая сохранность водорослей обеспечивается при
упаковке в водонепроницаемые бумажные пакеты и хранении в сухом помещении; в этих
условиях сырье с содержанием полисахаридов не менее 40% может храниться до 3 лет [5].
Выводы
1. Установлены основные морфологические признаки цельных слоевищ бурых
водорослей (ламинарии сахаристой, фукуса пузырчатого) и красной водоросли
(анфельции складчатой).
2. Выявлены диагностически значимые признаки измельченных слоевищ водорослей:
длинные трубчатые нити и слизистые каналы у ламинарии, наличие промежуточного слоя
у ламинарии и фукуса.
3. По товароведческим показателям, установленным для слоевищ ламинарии, фукус и
анфельция удовлетворяют требованиям ГФ ХI, кроме слоевищ анфельции по содержанию
песка.
4. Установлен идентичный качественный состав водорослей (полисахариды,
альгиновые кислоты, восстанавливающие сахара, йод, маннит, аминокислоты), кроме
наличия агара только в анфельции.
5. Определено количественное содержание действующих веществ в водорослях
(полисахариды, альгиновые кислоты, восстанавливающие сахара, пентозаны, йод,
клетчатка, маннит, белки, липиды, агар). В ламинарии, по сравнению с фукусом и
анфельцией, отмечена более высокая концентрация: альгиновых кислот (в 1,4 и 5,75 раз),
полисахаридов (в 1,3 и 1,4 раз), йода (в 4,5 и 1,8 раз), маннита (в 1,5 и 2,5 раз).
Максимальное накопление активных веществ в ламинарии наблюдается в слоевищах,
залегающих на глубине 10 м от поверхности моря, в период с сентября по октябрь.
6. Установлено активное накопление в водорослях натрия, калия, кальция, железа,
магния, марганца, обусловленное соответствующим минеральным составом морской
воды. Выведены математические уравнения регрессии между содержанием в водорослях
белков и марганца (уровень вероятности 99%), белков и железа (уровень вероятности
95%).
7. Естественные условия сушки и хранение сырья без доступа влаги обеспечивают
концентрацию полисахаридов не менее 40% в течение 3 лет.
Библиографический список
1. Антонова М.А. Методика количественного определения альгиновой кислоты //
Материалы рыбохоз. исследований Северного бассейна. – Мурманск, 1980. – С. 193–201.
2. ГОСТ 26185-84. – Введ. 1986.–07.01. Водоросли морские, травы морские и продукты
их переработки. Методы анализа: – М.: Изд-во стандартов, 1986. – 12 с.
Бурые водоросли — презентация онлайн
1. ДГТУ Кафедра ТСА Лабораторная работа № 7
• Отдел Отдел Бурые водоросли
• Число. Месяц.2013
• Выполнил
• Иванов А.А. гр. БПБ21
• Проверила
• доц. Нейдорф А.Р.
• Ростов-на-Дону
• 2013
2. Цель работы
• Ознакомиться с особенностями
организации бурых водорослей
3. ОТДЕЛ БУРЫЕ ВОДОРОСЛИ РНАЕОРНУТА
• морские, особенно много — в холодных водах северного и
южного полушария.
• неизвестны ни одноклеточные, ни колониальные формы,
ни талломы в виде простой неразветвленной нити.
• самые простейшие — гетеротрихальные
• талломы крупные, ложно- или истиннотканевого
строения.
4. ОТДЕЛ БУРЫЕ ВОДОРОСЛИ РНАЕОРНУТА
• КЛАСС ИЗОГЕНЕРАТНЫЕ
• КЛАСС ГЕТЕРОГЕНЕРАТНЫЕ
• КЛАСС ЦИКЛОСПОРОВЫЕ
4
6. КЛАСС ИЗОГЕНЕРАТНЫЕ
• Порядок эктокарповые
• род эктокарпус
— самые
примитивные бурые водоросли.
• Распространены во всех морях, особенно
холодных, и растут на подводных
предметах и других более крупных
водорослях.
6
7
8
9. КЛАСС ИЗОГЕНЕРАТНЫЕ
• характеризуются изоморфной сменой
генераций
• Изоморфная смена поколений — тип
жизненного цикла водорослей с наличием
диплоидной и гаплоидной фазы,
характеризующийся одинаковыми по
внешнему виду спорофитом и
гаметофитом.
10. КЛАСС ИЗОГЕНЕРАТНЫЕ
• Порядок эктокарповые
• род эктокарпус
— самые
примитивные бурые водоросли.
• Распространены во всех морях, особенно
холодных, и растут на подводных
предметах и других более крупных
водорослях.
10
11
12
13. Цикл развития Ectocarpus siliculosus
• На спорофите могут возникать как одногнездные,
так и многогнездные зооопорангии. В первых
послемейоза образуются гаплоидные n зооспоры,
во вторых — диплоидные (2n) зооспоры;
гаплоидные зооспоры прорастают в гаметофиты,
несущие многогнездные гаметангии . Гаметы
копулируют, зигота прорастает в диплоидный
спорофит. Диплоидные зооспоры сразу
прорастают с образованием диплоидного
спорофита
14. Порядок кутлериевые
• Порядок включает только два рода: кутлерия и
занардиния
• Кутлерия распространена по всему побережью
Европы, занардиния — преимущественно в
Средиземном море, а также в Черном.
• Оба рода обнаруживают чередование
поколений: у кутлерии смена генераций
гетероморфная, у занардинии — изоморфная
14
15. Рисунок 1. Гаметангии кутлерии
15
16
17. род падина
17
18. Рисунок 2. Padina pavonica
18
19. КЛАСС ГЕТЕРОГЕНЕРАТНЫЕ
• Гетероморфная смен поколений – тип
жизненного цикла водорослей с наличием
диплоидной и гаплоидной фазы,
характеризующийся резко отличающимися
по внешнему виду спорофитом и
гаметофитом. Редукционное деление ядра,
проходящее после некоторого времени
существования диплоидного таллома,
приводит к образованию не гамет, а спор
(спорическая редукция).
19
• Каждая из спор дает начало новому
растению, клетки которого оказываются
гаплоидными. В дальнейшем на таком
растении – уже без редукционного деления
– образуются гаметы, слияние которых
вновь приводит к образованию
диплоидного организма.
21. Порядок ламинариевые
• Гаметофиты мало различаются между
собой и представлены микроскопическими,
часто редуцированными до нескольких
клеток нитчатыми растеньицамизаростками, которые несут половые органы.
На мужских гаметофитах образуются
антеридии в виде мелких клеток,
развивающих по одному сперматозоиду, на
женских — оогонии, в которых
формируется по одной яйцеклетке.
22. Спорофиты
• разных родов резко различаются и представляют собой
самые крупные низшие растения, достигающие в длину
60—100 м,
• со значительным морфологическим расчленением и
сложным анатомическим строением. В большинстве
случаев спорофиты ламинариевых
• расчленяются на листовидную пластинку, «ствол» и
ризоиды, с помощью которых все растение прикрепляется
к подводным камням и скалам. В месте перехода
листовой пластинки в стволик находится
интеркалярная меристема, за счет деятельности
которой нарастает как листовая пластинка, так и
ствол.
• род ламинария, виды которого широко распространены в
северных морях.
23. Рисунок 2. Жизненный цикл ламинарии
24. КЛАСС ЦИКЛОСПОРОВЫЕ
Порядок фукусовые
• Характеризуется:
–
–
–
–
верхушечным ростом таллома
Отсутствием бесполого размножения
оогамным половым процессом
половые органы располагаются в углублениях таллома
— концептакулах, или скафидиях
• Имеется вегетативное размножение, приводящее в некоторых
участках океана к образованию громадных скоплений талломов
саргассума (примером может служить Саргассово море).
25. Рисунок 3 Зрелые концептакулы фукуса пузырчатого (Fucus vesiculosus): 1 — женский; 2 — мужской
26. Вывод
• Отдел бурые водоросли включает почти
исключительно морские водоросли,
• характерен особый набор пигментов,
• в основе классификации лежат
особенности жизненного цикла.
27. Вопросы для подготовки. Заполните таблицу
Класс бурых
водоролей
Класс
изогенератные
Класс
гетерогенератн
ые
Класс
циклоспоровые
Строение Особеннос распростран Представит
таллома
ти
ение
ели
жизненного
цикла
28. Темы для сообщений
1. Красные водоросли. Их систематика
2. Класс бангиевые. Особенности
строения и жизненные циклы
3. Класс флоридеи. Особенности
строения и жизненные циклы
Использование фукуса пузырчатого в овощеводстве открытого грунта Текст научной статьи по специальности «Сельское хозяйство, лесное хозяйство, рыбное хозяйство»
УДК:631.873:635.07
ИСПОЛЬЗОВАНИЕ ФУКУСА ПУЗЫРЧАТОГО В ОВОЩЕВОДСТВЕ ОТКРЫТОГО ГРУНТА
М.Г. Юркевич, к.с.-х.н. (рецензентМ.В. Медведева, к.б.н.)
Институт биологии Карельского научного центра РАН, e-mail: [email protected]
В статье рассмотрено использование водного настоя бурой морской водоросли фукус пузырчатый (Fucus vesiculosus L.) при выращивании огурцов в условиях открытого грунта. Установлено, что опрыскивание растений существенно (в 2 раза) повышает урожайность, а концентрация препарата в 100 г/л не вызывает ожогов листьев и требует небольшого количества сырья.
Ключевые слова: полевой опыт, водоросли, фукус, огурец, овощеводство, урожайность.
USE FUCUS VESICULOSUS IN VEGETABLE OPEN GROUND
M.G. Yurkevich
Institute of Biology, Karelian Research Centre RAS, e-mail: [email protected]
In the article the use of water extract of brown algae bladder wrack (Fucus vesiculosus L.) in plants cultivation of cucumbers in conditions of open ground. It is established that the spraying ofplants significantly (2 times) increases the yield and concentration of the drug in 100 g/l doesn’t cause burns leaves.
Keywords: field experiment, algae, fucus, cucumber, vegetable, yields.
Россия обладает огромным ресурсным потенциалом морских водорослей. Запас только основных промысловых и потенциально промысловых видов (фукусовые, ламинарии и алярия) составляет более 13,5-30,0 млн. т сырой массы [1, 2], а общий рекомендованный объем добычи всех видов водорослей составляет почти 230 тыс. т [3]. Однако по целому ряду экономических причин освоение этого объема осуществляется не более чем на 30% [4].
Фукус пузырчатый (Fucus vesiculosus L.) — один из основных промысловых видов морских бурых водорослей. Этот бореально-арктический вид распространен в Атлантическом и Северном Ледовитом океанах является массовым видом литорали Баренцева и Белого морей. Так, на мурманском побережье Баренцева моря запас фукусовых водорослей составляет около 180 тыс. т, в Белом море общий запас литоральных фукоидов оценивается примерно в 250 тыс. т, а на Российском побережье Балтики составляет 75 тыс. т [3]. В настоящее время отсутствует промышленный промысел морских водорослей на побережье Баренцева и Балтийского морей. На побережье Белого моря предприятиями Архангельской области ежегодно добывается не более 0,3-0,5 тыс. т фукусовых водорослей. В республике Карелия ежегодная квота добычи беломорского фукуса составляет 1600 т, однако ее практически не выбирают [5]. Поэтому у приморских регионов есть существенный потенциал в расширении и восстановлении этой сферы добычи переработки морских ресурсов.
Исследования по использованию фукуса ориентированы на пищевую и косметическую промышленность, в меньшей степени — на медицинские исследования. Разработаны технологии применения фукуса в производстве кондитерских изделий, пищевых биодобавок, косметических кремов, масок и т.д.
В сельском хозяйстве фукус в качестве кормовой добавки нашел применение в птицеводстве [5, 6]. В ряде
работ [7, 8] показана высокая эффективность предпосевной обработки семян сои препаратами на основе фукуса: урожайность культуры повышается на 3,1-3,9 ц/га (15,6%), размеры корней увеличиваются на 20%, стебля — на 26%, также сокращается продолжительность вегетационного периода на 3-4 дня. Однако все разработанные технологии использования бурых водорослей требуют либо специального оборудования, либо основаны на применении сложных по производству экстрактов водорослей. По этой причине они не получили массового распространения.
Целью исследований было предложить малозатратную технологию использования фукуса в овощеводстве, доступную всем категориям сельскохозяйственных производителей. Для этого был проведен полевой эксперимент по изучению влияния водного настоя фукуса пузырчатого на рост и развитие растений огурца в открытом грунте.
Эксперимент проводили на базе Корзинского научного стационара Института биологии КарНЦ РАН (южная Карелия). Опыт был заложен посевом семян огурца сорта Кураж в открытый грунт. Площадь опытной делянки составляла 2 м2, в каждом варианте было по три растения, повторность выполнения опыта — трехкратная. Почвы опытного участка супесчаные, слабокислые (рНкс1 5,24), характеризуются низким содержанием калия (52 мг/кг) и фосфора (41,8 мг/кг). В эксперименте исследовали вариант без применения настоя фукуса (контроль) и три варианта с применением водного настоя из сухой крупки фукуса пузырчатого в концентрациях 150 г, 100 г и 50 г крупки на 1 л воды, на каждое опытное растение использовали 1 л настоя.
В течение вегетационного сезона проводили подкормку растений минеральными удобрениями: двукратно N30 и трехкратно Р2оК20. В течение сезона было проведено два плановых опрыскивания растений настоем фукуса: в фазе массового цветения и в период массового
Биометрические показатели
развития растении огурца
и урожайность
Вариант Количество Количество Длина основ- Урожайность,
стеблей, шт. листьев, шт. ного побега, см кг/м2
Контроль (без опрыскивания) 7,63±2,35 74,50±33,93 133,75±34,63 1,67
Опрыскивание растений настоем фукуса, 150 г/л 8,56±1,20 86,56±17,43 178,89±30,00 4,00
Опрыскивание растений настоем фукуса, 100 г/л 9,11±1,27 96,22±22,67 188,89±19,64 3,79
Опрыскивание растений настоем фукуса, 50 г/л 9,33±1,55 88,00±12,72 176,11±20,95 2,08
600 500 £ 400
о
0 и
300
1
^ 200 100 0
06.08. 09.08. 12.08. 15.08. 19.08. 22.08. 26.08. 29.08. 03.09.
сроки сбора урожая -контроль — — 2 вариант — — — 3 вариант — — — ■ 4 вариант
г/м2
2
Урожайность растений огурца, г/м
начала плодоношения. Урожай собирали с периодичностью 1 раз в три-четыре дня. Первый сбор урожая был 06.08.2013, последний — 03.09.2013. Средний размер плода составил 12 см. Статистическую обработку данных проводили в программе Statgraphis plus и по методике Доспехова (1985).
При проведении эксперимента оценивали биометрические показатели развития растений огурца. Установлено, что применение настоя фукуса не оказывает существенного влияния на количество побегов и листьев, а также на длину основного стебля (таблица).
Анализ устойчивости растений к заболеваниям показал, что в течение вегетационного периода растения огурца контрольного варианта (от 4 до 11%) были подвержены заражению фузариозным и бактериальным увяданием. В тоже время растения, обработанные настоем фукуса в концентрации 100 г/л и 50 г/л, в течение всего сезона были здоровы (0-2% пораженных листьев от общего количества), однако при опрыскивании мак-
симальной концентрацией (150 г/л) были отмечены признаки ожога листа (4-6% пораженных листьев).
Суммарно за вегетационный сезон урожайность огурца достоверно отличалась в зависимости от концентрации настоя фукуса при опрыскивании, предел ее варьирования составил от 1,67 кг/м2 в контроле до 4,00 кг/м2 в варианте с максимальной концентрацией настоя фукуса (таблица).
При периодических сборах продуктивность растений в контроле и с применением минимальной концентрации фукуса (вариант 4) была существенно ниже, чем продуктивность растений при опрыскивании средними и высокими концентрациями — варианты 2 и 3 (рисунок).
Таким образом, при опрыскивании растений огурца водным настоем фукуса пузырчатого оптимальная доза составила 100 г/л. При этом существенно увеличилась продуктивность (в 2,3 раза). Данная концентрация также не вызывает ожогов листьев и требует меньшего количества сырья.
Литература
1. Уховеева М.В., Подкорытова А.В. Промысловые водоросли и травы морей Дальнего Востока: биология, распространение, запасы, технология переработки. — Владивосток: ТИНРО-Центр, 2006. — 243 с.
2. Евсеева Н.В. Макрофитобентос прибрежной зоны Южных Курильских островов: состав, распределение и ресурсы: автореф. дисс. к.б.н. — М., 2009. — 22 с.
3. Вилкова О.Ю. Место России в мировой добыче морских водорослей // Рыбпром: Технологии и оборудование для переработки водных биоресурсов, 2010, № 3. — С. 4-8.
4. Food and Agricultural Organization of the United Nations, 2008. «Fishery and Aquaculture Statistics».
5. Шарапова В.Ю. Беломорские фукусы в рационах птицы // Птицеводство, 2011, № 9. — С. 23-24.
6. Величко О.А. Морские водоросли фукус в рационах кур-несушек // Птица и птицепродукты, 2010, № 5. — С. 50.
7. Заостровных В.И., Симкина Т.Ф., Шевченко Н.М., Чайкина Е.Л., Звягинцева Т.Н., Анисимов М.М. Продуктивность сои и ее устойчивость к заболеваниям под влиянием углеводсодержащих биополимеров из морских водорослей // Достижения науки и техники АПК, 2010, № 5. — С. 42-43.
8. Имбс Т.И., Чайкина Е.Л., Дега Л.А., Ващенко А.П., Анисимов М.М. Сравнительное изучение химического состава этанольных экстрактов бурых водорослей и их влияния на рост проростков и урожайность сои Glycine max (L). Merr // Химия растительного сырья, 2010, № 1. — С. 143-148.
|
Маникюр мужской |
Продолжительность |
Стоимость |
|---|---|---|
|
Обрезной маникюр |
45 мин |
850 |
|
Европейский маникюр (необрезной) |
45 мин |
850 |
|
Аппаратный маникюр |
45 мин |
850 |
|
Комбинированный маникюр |
45 мин |
850 |
|
SPA-маникюр (с обработкой кутикулы) |
70 мин |
1 400 |
|
Полировка шлифовка |
10 мин |
150 |
|
Придание формы |
10 мин |
200 |
|
Нанесение крема |
10 мин |
150 |
|
Педикюр мужской |
Продолжительность |
Стоимость |
|
Обрезной педикюр |
60 мин |
1 600 |
|
Европейский педикюр |
60 мин |
1 600 |
|
Аппаратный педикюр |
60 мин |
1 600 |
|
Комбинированный педикюр |
60 мин |
1 600 |
|
SPA-педикюр (с обработкой стоп) |
75 мин |
2 300 |
|
SPA-уход за руками |
Продолжительность |
Стоимость |
|
«Бархатный комплекс» (ванночка для рук, пилинг, маска, массаж, крем) |
40 мин |
700 |
|
Парафиновая ванночка для рук |
30 мин |
400 |
|
SPA-уход за ногами |
Продолжительность |
Стоимость |
|
«Бархатный комплекс» (ванночка для ног, пилинг, маска, массаж, крем) |
40 мин |
1 100 |
|
Покрытие ногтей |
Продолжительность |
Стоимость |
|
Снятие лака |
10 мин |
100 |
|
Покрытие ногтей лаком |
15 мин |
250 |
|
Покрытие лаком French |
20 мин |
350 |
|
Покрытие ногтей лечебное |
10 мин |
200 |
|
Питание ногтевой пластины (пчелиный воск с шёлком) |
20 мин |
600 |
|
Покрытие лечебное защитное (перманентное глянцевание) |
30 мин |
900 |
|
Покрытие «Быстрая сушка» |
10 мин |
100 |
|
Снятие гель-лака/Shellac |
20 мин |
400 |
|
Маникюр с покрытием Shellac |
30 мин |
1 000 |
|
Маникюр с покрытием French Shellac |
40 мин |
1 200 |
|
Педикюр с покрытием Shellac |
30 мин |
1 100 |
|
Педикюр с покрытием French Shellac |
30 мин |
1 300 |
|
Дизайн ногтя (рисунок) |
15 мин |
100 |
|
Дизайн ногтя (стразы) |
5 мин |
50 |
|
Депиляция восковая |
Продолжительность |
Стоимость |
|
Щёки |
15 мин |
300 |
|
Верхняя губа |
15 мин |
300 |
|
Подмышечные впадины |
15 мин |
600 |
|
Руки полностью |
25 мин |
1 000 |
|
Руки до локтя |
15 мин |
600 |
|
Кисти рук |
15 мин |
400 |
|
Линия живота |
10 мин |
400 |
|
Живот |
15 мин |
900 |
|
Ягодицы |
20 мин |
1 000 |
|
Бикини классическое |
30 мин |
1 000 |
|
Бикини глубокое |
35 мин |
1 500 |
|
Ноги полностью |
30 мин |
1 800 |
|
Бедра |
20 мин |
1 000 |
|
Голень (до колена) |
30 мин |
1 200 |
|
Пальцы ног |
10 мин |
300 |
|
Пилинг |
Продолжительность |
Стоимость |
|
Пилинг с солевым скрабом |
30 мин |
2 000 |
|
Пилинг рукавичкой «Кесе» |
30 мин |
1 500 |
|
Пилинг «Морской бриз» (морская соль, водоросли, экстракты трав) |
40 мин |
2 000 |
|
Турецкий мыльный массаж (процедура, завершающая пилинг) |
30 мин |
1 800 |
|
Обертывания |
Продолжительность |
Стоимость |
|
Обёртывание «Солевое» |
60 мин |
3 200 |
|
Обёртывание «Масленое» |
60 мин |
3 700 |
|
Обёртывание «Морской гель» |
60 мин |
3 300 |
|
Обертывание «Морские водоросли» |
60 мин |
2 600 |
|
Обертывание «Шоколадное» |
60 мин |
2 600 |
|
SPA-программы для тела |
Продолжительность |
Стоимость |
|
«Зимняя вишня». Программа с холодным обёртыванием (пилинг с морской солью, фукус, ламинария, холодное водорослевое обёртывание, питание водорослевым маслом) |
90 мин |
4 800 |
|
«Жаркие объятия». Антицеллюлитная с горчицей и мёдом (пилинг с морской солью, фукус, ламинария, горячее водорослевое обёртывание (горчица+мёд), питание SLIM- маслом) |
90 мин |
4 800 |
|
«Морской бриз». Комплексный уход для тела (пилинг с морской солью, фукус, ламинария, водорослевое обёртывание, питание водорослевым маслом) |
90 мин |
5 000 |
|
«Анти-Эйдж». Омолаживающая SPA-программа для тела (пилинг с морской солью, фукус, ламинария, комбинированное водорослевое обёртывание питание SLIM-маслом, интенсивное увлажнение) |
90 мин |
5 500 |
|
SPA-программы для тела |
Продолжительность |
Стоимость |
|
«Магия красоты». Антицеллюлитная программа (пилинг морской солью, фукус, ламинария, водорослевое обёртывание питание SLIM- маслом, экстракт фукуса) |
90 мин |
4 800 |
|
«Идеальный силуэт». Программа для похудения (пилинг морской солью, фукус, ламинария, водорослевое обёртывание питание липолитической сывороткой) |
90 мин |
5 000 |
|
«Морская жемчужина». Лимфодренажная про- грамма (пилинг морской солью, фукус, ламинария, ком- бинированное водорослевое обёртывание, питание SLIM-маслом, экстракт ламинарии) |
90 мин |
4 800 |
|
SPA-программы для ног |
Продолжительность |
Стоимость |
|
«SPA-уход для ног» |
50 мин |
2 400 |
|
«Здоровые ножки» | 50 мин |
2 400 |
|
«Легкие ножки» (противоварикозная) |
50 мин |
2 400 |
|
«Стройные ножки» (против отёков) |
50 мин |
2 400 |
|
«Экспресс-уход для ног» |
50 мин |
1 200 |
|
SPA-программа для женщин |
Продолжительность |
Стоимость |
|
«Привлекательная и обаятельная». Антицеллюлитная, проблемные зоны (пилинг морской солью, фукус, ламинария, комбинированное водорослевое обёртывание, питание липолитической сывороткой и SLIM-маслом) |
75 мин |
3 500 |
|
«Пьянящий шоколад». Антицеллюлитная (на выбор распаривание в инфракрасной сауне или антицеллюлитный массаж, шоколадный пилинг тела, шоколадное обертывание, нанесение увлажняющего крема для тела, чайная церемония) |
120 мин |
6 000 |
|
SPA-ванна |
Продолжительность |
Стоимость |
|
Молочная ванна «Клеопатра». Комплесный уход за кожей. Воздействие активных компонентов направлено на омоложение, снятие стресса и релаксирующее действие на кожу |
60 мин |
2 800 |
|
SPA-программа для мужчин |
Продолжительность |
Стоимость |
|
«Время расслабиться!». В стоимость входит: парение одного человека в сауне «Лагуна», прогрев и парение вениками в 2 захода, медово-солевой пилинг, СПА-массаж «Лимфодренажный», посещение купели, чайная церемония |
2 часа |
6 000 |
Развитие бурой водоросли Fucus evanescens Ag. в прикамчатских водах + «
Автореферат диссертации по теме «Развитие бурой водоросли Fucus evanescens Ag. в прикамчатских водах»
На правах рукописи
ЧМЫХАЛОВА Виктория Борисовна
РАЗВИТИЕ БУРОЙ ВОДОРОСЛИ FUCUS EVANESCENS Ag. В ПРИКАМЧАТСКИХ ВОДАХ
03.00.18 — Гидробиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени кандидата биологических наук
Петропавловск-Камчатский 2005
Работа выполнена в Камчатском государственном техническом университете, г. Петропавловск-Камчатский
Научный руководитель
доктор биологических наук Н. часов, на заседании диссертационного совета КМ 307. 008.01 по защите диссертаций на соискание ученой степени кандидата биологических наук при Камчатском государственном техническом университете.
Отзывы на автореферат диссертации в двух экземплярах с заверенными подписями просим направлять по адресу: 683003, г. Петропавловск-Камчатский, ул. Ключевская, 35, Камчатский государственный технический унйверситет (КамчатГТУ), диссертационный совет КМ 307. 008.01.
Телефон: (4152) 12-45-38. Факс: (415-2) 12-05-01.
‘ I
С диссертацией можно ознакомиться в научной библиотеке Камчатского государственного технического университета.
Автореферат разослан « 3 » октября 2005 г.
Ученый секретарь диссертационного совета( к. б. и.
Л.В. Ромейко
\
-ч
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследований. Порядок Fucales — один из крупнейших среди бурых водорослей. Он представлен во флоре Мирового океана большим числом семейств и родов. Интерес к его изучению обусловлен значительной ролью его представителей в формировании структуры растительности и условий обитания прибрежных гидробионтов. Практическая значимость фукусовых определяется их ценным химическим составом.
Среди родов, встречающихся в холодоумеренных водах Северного полушария, наиболее широкое распространение имеет род Fucus. Он же является здесь самым многочисленным. В прикамчатских водах встречается единственный представитель этого рода F. evanescens Ag. Его массовое развитие и практически повсеместное распространение в литоральной зоне шельфа позволяют рассматривать этот вид как потенциально промысловый. Рациональное использование ресурсов фукуса и фукусового сырья должно основываться на данных изучения его роста и развития, химического состава, структуры популяций, влияния на них условий обитания.
В последние годы прибрежные воды камчатского шельфа подвергаются антропогенному загрязнению. Для оценки экологического состояния вод наряду с гидрохимическими могут использоваться гидробиологические показатели, в частности показатели макрофитобентоса на всех уровнях его организации (Клочкова, 1998; Клочкова, Березовкая, 2001; Березовская, 2002). Все виды рода Fucus демонстрируют высокую экологическую пластичность, способны выносить высокий уровень загрязнения (Христофорова, 1981; 1995; и др.). Многолетний цикл жизни, простота определения возраста делают их удобными для проведения экологического мониторинга в прибрежных акваториях.
Представитель камчатской флоры F. evanescens благодаря хорошо выраженной, как и у других видов рода Fucus, возрастной, сезонной, географической и экологической изменчивости привлекателен как модельный объект для изучения ответных реакций организмов и популяций на воздействие неблагоприятных факторов. Ответный отклик на организменном уровне надежнее и зримее всего проявляется в изменении морфометрических параметров, морфо-физиологического состояния, темпов роста и развития, а в популяционном -на изменении возрастной структуры популяций и адаптивной стратегии размножения. i
Определяя тему исследования, мы полагали, что изучение этих вопросов для камчатской популяции фукуса позволит не толькр заложить научный фундамент для разработки вопросов рационального использования этого вида, но также даст надежную основу для использования его в качестве объекта биомониторинга состояния литоральных сообществ бентоса.ювнях для последующего практического использования х водах.
Для достижения поставленной цели необходимо было решить следующие задачи:
— изучить распространение рода Fucus, особенности биологии развития его представителей, их ответные реакции на факторы среды в других районах Мирового океана для выбора районов проведения регулярных наблюдений;
— изучить особенности морфогенеза растений на первом году жизни и на этой основе выбрать методику определения их возраста, установить продолжительность жизни фукуса у камчатских популяций вида;
— изучить особенности сезонного линейного роста у разновозрастных представителей популяции в разных условиях обитания и определить воздействие на них загрязнения и основных гидрологических факторов;
— определить сезонную динамику изменения массы у разновозрастных растений, обитающих в разных условиях обитания;
— выявить изменение стратегии размножения вида под воздействием неблагоприятных факторов;
— выявить сезонные изменения и особенности формирования возрастной структуры популяций, стратегию ее изменения под воздействием неблагоприятных факторов;
— определить функциональную роль разных частей слоевища по содержанию в них пластических веществ, сезонные и возрастные изменения их содержания и причины, влияющие на эти изменения;
— выявить основные направления адаптивных изменений организма и популяции к стрессовым нагрузкам среды;
— разработать рекомендации по рациональному использованию фукусового сырья в прикамчатских водах.
Научная новизна. Впервые у представителей вида камчатской популяции F. evanescens установлена продолжительность жизни в зависимости от условий обитания и выявлена стратегия развития популяций. Определена роль разных генераций в воспроизводстве, формировании основной продукции, ее сезонном изменении. Выявлены адаптационные приспособления к неблагоприятным факторам природной среды и к стрессовому воздействию загрязнения на организменном и популяционном уровнях.
В каждый период жизни растений выявлены характеристики морфо-физиологического состояния, которые можно с успехом применять как показатели экологического состояния среды обитания.
Впервые у каждой возрастной генерации вида изучены сезонные изменения содержания сухих веществ в разных частях слоевища, установлена зависимость этих изменений от состояния среды обитания и показано, что характер этих изменений хорошо отражает физиологическое состояние растений.
Практическое значение. Результаты исследований могут бьггь использованы для разработки рекомендаций по промыслу F. evanescens, поскольку позволяют установить сроки промысла и нормы изъятия водорослей. Данные по содержанию у F. evanescens пластических веществ позволяют определить направления использования сырья. Данные по развитию фукуса в антропогенной
среде могут быть использованы для оценки экологического состояния прибрежных вод, где этот вид имеет массовое распространение.
Апробация работы. Основные результаты исследований докладывались и представлялись на научно-техническом симпозиуме «Современные средства воспроизводства и использование водных биоресурсов» (Санкт-Петербург, 2000), научно-технических конференциях профессорско-преподавательского состава КамчатГТУ (Петропавловск-Камчатский, 2001; 2002; 2003; 2004 и 2005), II, III и V научных конференциях «Сохранение биоразнообразия Камчатки и прилегающих морей» (Петропавловск-Камчатский, 2001; 2002; 2004), научно-технической конференции «Рациональное использование морских биоресурсов» (Петропавловск-Камчатский, 2002), научно-технической конференции «Экономические, социальные, правовые и экологические проблемы Охотского моря и пути их решения» (Петропавловск-Камчатский, 2004). Результаты работы докладывались также на заседаниях Камчатского отделения Всероссийского ботанического общества (КО РБО) (2002; 2003), а также на семинарах и расширенных заседаниях лаборатории альгологии Камчатского филиала тихоокеанского института географии ДВО РАН.
Публикации. Материалы диссертации представлены в 11 публикациях.
Структура и объем диссертации. Диссертация состоит из введения, 8 глав, заключения, выводов и списка литературы, включающего 144 источника, из них 38 на иностранном языке. Работа изложена на 137 страницах, иллюстрирована 42 рисунками и 16 таблицами.
Благодарность. Выражаю искреннюю признательность моему научному руководителю д.б.н. Н.Г. Клочковой. Я также весьма признательна заведующей кафедрой технологии рыбных продуктов КамчатГТУ М.В. Ефимовой за постоянную помощь и поддержку при написании диссертации, сотрудникам лаборатории КФ ТИГ ДВО РАН к.б.н. Т.Н. Королевой и м.н.с. А.Э. Кусиди за помощь в сборе материала и выполнении работы на всех ее этапах. Благодарю д.г.н. В.А. Березовскую за консультации по вопросам антропогенного загрязнения Авачинской губы. Признательна декану технологического факультета КамчатГТУ к.т.н. В.Н. Дегтяреву за моральную поддержку.
СОДЕРЖАНИЕ РАБОТЫ ГЛАВА 1. ЛИТЕРАТУРНЫЙ ОБЗОР
Фукусовые водоросли всегда являлись объектом пристального внимания исследователей, и их изучение проводилось в самых разных направлениях. В главе дается обзор публикаций, касающихся представителей порядка Fucales и рода Fucus. Приводятся данные по состоянию изученности представителей рода, обитающих у арктического, атлантического и тихоокеанского побережий, а также рассматриваются работы, в которых приводятся любые упоминания о F. evanescens на российском Дальнем Востоке. Показано, что опубликованные
к настоящему времени работы содержат в основном сведения о нахождении вида в разных районах побережья или краткие сведения о составе и биомассе фукусовых сообществ.
Анализ альгологических и гидробиологических работ показал, что, несмотря на многочисленные упоминания вида в научной литературе по Дальнему Востоку, развитие Р. еуапеясепя здесь практически не изучено. Большинство данных по разным вопросам роста, развития и размножения фукусов было получено для беломоро-балтийских популяций из ненарушенных условий произрастания. Разностороннее изучение влияния антропогенного загрязнения на развитие и воспроизводство популяций фукуса и стратегию их адаптивных изменений до сих пор не проводилось.
ГЛАВА 2. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Материалом исследований являлись образцы водорослей, собранных у берегов юго-восточной Камчатки. Привлекались также сборы и гербарные материалы по фукусам других районов обитания Дальнего Востока: Командорских и северных Курильских островов, северо-восточной и западной Камчатки и др.
Большая часть материала была собрана в период с 1998 по 2002 год в Авачинском заливе и представляла собой сезонные сборы. Для изучения развития растений в природной среде была выбрана б. Вилючинская. Изучение влияния загрязнения на развитие растений проводилось в Авачинской губе, характеризующейся высоким уровнем антропогенного загрязнения. О характере загрязнения разных прибрежных участков этого водоема судили по данным публикаций (Березовская, 1988, 2002; Клочкова, Березовская, 2001, и др.). На основе этих данных были выбраны три полигона с разным типом и уровнем антропогенного воздействия: в б. Моховой, прибрежной зоне в районе судоремонтного завода (СРМЗ) и б. Сероглазка. С 1998 по 2000 год на выбранных участках производили ежемесячный (с мая по ноябрь) отбор проб. В зимний и ранневесенний период из-за мощного ледового припая и большого снежного покрова сбор материала не производили. В 2001-2002 годах отбор проб проводился один раз в сезон. В необходимых случаях брались единичные пробы. В 2003 году было выполнено однократное обзорное исследование состояния популяций фукуса в июне в период большого суточного отлива.
Для сбора количественных проб на каждом из полигонов выбирались хорошо опознаваемые участки с разновозрастными зарослями. Длина таких участков составляла 30-50 м. Для выбора методики определения возраста и годового прироста растений в первый год исследований на часть растущих растений разного возраста были установлены метки. Для этого стебель растения оборачивали полипропиленовой лентой и скрепляли ее края степлером так, чтобы она достаточно жестко крепилась к растению. На ленте водостойким маркером проставляли номер образца. В 1998, 1999 и 2000 годах таким способом было помечено 15-20 растений каждой возрастной группы на каждом участке каждо-
го полигона. Для удобства наблюдений для каждого года использовали ленты разного цвета.
В ходе полевых исследований меченые образцы просматривались каждый месяц для определения особенностей развития дихотомических ветвей, времени вступления растений в размножение, а также для определения сроков закладки, созревания, сброса рецептакулов у растений разных возрастов. Всего за период исследований было помечено 897 особей, из которых до конца наблюдений дожили 586.
Пробы фукуса для лабораторных исследований собирали во время сиги-зийных отливов в среднем горизонте литорали. Для взятия количественных проб использовали рамку 0,25×0,25 м2. Ее накладывали на участок дна с наиболее высокой для района плотностью зарослей, а затем выбирали из нее все экземпляры, подошва которых приходилась на площадь, ограниченную рамкой. Одна проба обычно включала 95-105 особей. Сбор проб на разных полигонах производили либо в один и тот же день, либо в течение двух дней. Период между помесячным отбором проб составлял обычно 26-30 дней.
В ходе обработки проб собранные растения разбирали по возрастам и определяли численный состав каждой возрастной группы. Далее в группах, состоящих из большого числа особей, отбирались для измерений по 30 растений. В группах, состоящих из меньшего числа особей, измерению подвергались все экземпляры. В ходе обработки проб у изучаемых растений определяли общую длину, массу, состояние фертильности, количество дихотомических разветвлений, количество ветвей, прекративших рост. Все полученные данные были занесены в электронную базу данных. Их обработка была выполнена с помощью компьютерной программы «Microsoft Excel-2000». Общее количество измеренных экземпляров за весь период исследования составило около 17 тыс. При этом в Авачинской губе было изучено более 14 тыс. образцов.
Возраст растений определяли по регистрирующему признаку — количеству дихотомических ветвлений слоевища. Исследования ряда авторов показывают, что у некоторых видов рода фукус в течение года может появляться несколько ди- или трихотомий (Максимова, 1980; Возжинская, 1986). Для получения статистически достоверных данных по возрастному развитию F. evanes-cens мы определяли, сколько ди- или трихотомических ветвей в течение года образуются у камчатских представителей вида.
Проведенные наблюдения показали, что морфогенез F. evanescens может идти по двум сценариям (рис. 1). Согласно первому проростки первого года жизни к зиме вмерзают в лед, не имея дихотомическое ветвления. В этом случае первая дихотомия у них образуется лишь в следующем вегетационном периоде. Далее каждый год у растений появляется только одно новое дихотомическое разветвление. В ходе такого развития растений количество дихотомий у них на единицу меньше, чем их возраст. Однако случаи проявления такого сценария развития составили не более 0,5 % всех изученных образцов.
Рис. 1. Возрастные изменения морфологии фукусов при разных типах первогодних проростков
Согласно второму сценарию морфогенеза к концу первого года жизни у проростков фукуса уже образуется заметно выраженная дихотомия, поэтому количество дихотомий у сформировавшихся из них растений строго соответствует их возрасту. Поскольку у камчатских представителей фукуса явно доминировал второй сценарий развития, он и был принят в нашей работе за основу методики определения их возраста.
Определение содержания воды и сухого вещества в разных частях слоевища проводили по стандартной методике по ГОСТ 7636. При определении этих показателей для разных частей слоевища составляли средние пробы из фрагментов, взятых не менее чем у 25-30 растений.
ГЛАВА 3. ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
В главе приводится краткая физико-географическая характеристика при-камчатских вод, описываются климатические и гидрологические особенности отдельных участков побережья, в которых был собран изучаемый нами материал. Показано, что Авачинская губа и б. Вилючинская, представляющие собой глубоководные бухты фиордового типа, достаточно близки по геоморфологическим и гидрологическим особенностям. Подробно рассмотрены сезонные изменения температуры, концентрации биогенов, ритмика приливно-отливных колебаний у юго-восточной Камчатки. Расчетным путем выявлены периоды времени, в течение которых фукус подвергается длительному иссушению. В течение лета наблюдается несколько таких периодов, которые могут длиться до 20 часов при постоянном солнечном освещении и влияют на ростовые процессы и сроки вегетации. Подробно описана ледовая обстановка, поскольку она во многом определяет особенности развития вида. Несмотря на географическую приближенность между собой полигонов исследований, перечисленные характеристики имеют в них свои особенности. Они помогают проследить, каким образом фукус реагирует на стрессовые условия.
ГЛАВА 4. ОБЩАЯ ХАРАКТЕРИСТИКА МОРФОЛОГО-АНАТОМИЧЕСКОЙ ОРГАНИЗАЦИИ, ВОСПРОИЗВОДСТВА ПРЕДСТАВИТЕЛЕЙ РОДА FUCUS И ВИДА F. EVANESCENS
I
В главе приводится характеристика морфолого-анатомической организации представителей рода Fucus и вида F. evanescens в частности. Подробно охарактеризованы процессы размножения фукусов, рассмотрены особенности сезонного развития растений, описано влияние факторов среды — температуры, освещенности, длительного иссушения и ледового покрова на развитие фукусов. Ростовые процессы и химический состав растений сильно изменяются в зависимости от перечисленных факторов.
Проведенные исследования показывают, что в Авачинской губе с конца ноября по середину апреля растения полностью вмерзают в лед и в таком состоянии существуют несколько месяцев. Рост фукуса начинается весной, после таяния льда. В этот период граница между темными прошлогодними и молодыми светло-оливковыми участками талломов становится отчетливой. Особенно хорошо рост выражен у мелких, до 3 см высоты, растений.
Проведенные исследования позволили говорить о том, что последующее увеличение фотосинтетически активной радиации (ФАР) и повышение температуры воды до 5-7°С приводит к массовому весеннему развитию всей популяции фукусов. При этом прирост растений резко увеличивается, возрастает их масса, закладываются органы размножения. С подогревом воды до 8 С начинается созревание рецептакулов. Дальнейшее повышение температуры воды до 10-15°С активизируют процессы размножения фукуса. Температурный оптимум размножения для фукуса составляет 10-17°С. Дальнейшее повышение температуры приводит к торможению всех процессов и отрицательно влияет на состояние слоевища. В целом стрессовые колебания температурного фактора служат толчком для стимуляции разных процессов: роста, закладки фертильной ткани, созревания и высева продуктов размножения. Отмеченная в ходе изучения развития фукусов тенденция совпадает с результатами исследований В.Б. Возжинской (1980).
В середине лета линейный рост фукусов заметно приостанавливается. ФАР в это время достаточно высока (3-10 ккал/см2/сут), температура воды благоприятна для развития водорослей. Однако в прибрежных водах умеренных широт, в том числе и у побережья Камчатки, наблюдается дефицит биогенов, главным образом азотсодержащих соединений (Возжинская, 1986; Березовская, 1999).
Созревание рецептакулов завершается к концу лета — началу осени. Их окраска изменяется от оливково-зеленой до желтой. После выброса из скафиди-ев продуктов размножения рецептакулы темнеют, усыхают и опадают. Период сброса рецептакулов для популяции, судя по нашим наблюдениям, может продолжаться 10-14 дней в зависимости от динамики температурных изменений. Однако в некоторые аномально теплые годы, каким, например, был 1999 год, сброс рецептакулов может происходить на одну-две недели раньше, чем обычно, и может длиться всего 2-3 дня. Интересно отметить, что одновременный сброс рецептакулов сопровождается отчетливо уловимым потрескиванием. Весь берег в эти дни бывает усыпан опавшими рецептакулами.
Завершают главу данные о распространении и ценотической роли F. evanescens на российском Дальнем Востоке и в прикамчатских водах. Эти данные позволяют говорить о том, что камчатская бурая водоросль F. evanescens как потенциально промысловый вид и индикатор состояния окружающей среды является одним из наиболее перспективных объектов научного исследования и методов оценки антропогенного загрязнения прибрежных вод, а также модельным объектом в экологических и экофизиологических исследованиях.
ГЛАВА 5. СЕЗОННЫЙ ЛИНЕЙНЫЙ РОСТ У FUCUS EVANESCENS В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ
В главе приведен краткий обзор результатов исследования динамики роста, развития и накопления массы К е\>апе$сет. Изучение роста и увеличение массы фукусов мы наблюдали в период 1998-2000 годов. В течение всего периода развития представители каждой возрастной генерации (1-7 годов жизни) из разных районов произрастания имеют свои особенности.
Особенности распределения средних показателей длины растений у каждой возрастной группы в июне и июле 1998 года показаны на рисунке 2 А.
Сравнение средней длины фукуса из разных районов исследования показывает, что в июне в каждой возрастной группе наибольшая длина отмечена у растений из бухты Вилючинской.
б Моховая
б Сероглазка
СРМЗ
Рис. 2. Особенности распределения средних показателей длины разновозрастных представителей Fucus çvanescens в июне (А) и июле (Б)1998 г.
При сравнении этих показателей у растений из внутренних районов Ава-чинской губы можно видеть, что здесь у всех возрастных групп самая большая длина наблюдается у растений в б. Моховая, а самая минимальная — в б. Сероглазка.
Пределы изменчивости средних показателей длины фукуса можно проследить, сравнив ее максимальное и минимальное значение в каждой возрастной группе. Сравнение приведенных в главе 5 относительных значений между собой показывает, что наиболее заметное отставание в росте демонстрируют одно-, двух- и шестилетние растения, а средние размеры семилетних фукусов в разных местах обитания выравниваются, и разница между максимальным и минимальным для этого возраста значениями длины сокращается до 17,5%.
Анализ размерных характеристик в июне у микропопуляций из разных мест обитания показывает, что различия в условиях обитания существенно влияют на скорость активного линейного роста. Размерная структура микропопуляций в каждом из районов имеет свои особенности, выраженные в том, что на момент окончания активного роста разница между длиной каждой предыдущей и последующих генераций в каждом районе имеет свойственные только ему значения.
К июлю (рис. 2 Б) средняя длина растений во всех возрастных группах, кроме растений первого года жизни, увеличилась, то есть их линейный рост стал менее интенсивным. Наши исследования показывают, что темпы месячного прироста у фукусов разных возрастов в указанных районах были неодинаковыми. В б. Моховая наиболее значительно выросли двух-, трех- и пятилетние растения, в районе СРМЗ — четырех- и семилетние.
В августе (рис. 3 А), как упоминалось выше, у возрастной группы растений, вступающей в размножение, в целом сформированы рецептакулы и в них идет активное созревание гамет и их подготовка к оплодотворению. Длина фукусов, как это видно из рисунка 3 А, с июля по август продолжала увеличиваться. Ее прирост оказался максимальным у фукусов из б. Вилючинская. В Ава-чинской губе среди однолетних растений наибольшую длину имели водоросли из б. Моховая. В районе СРМЗ они дали’ незначительный прирост, в б. Серо-глазка — минимальный. В б. Моховая наиболее активно по-прежнему росли трех-, четырех- и семилетние растения.□4 года | Н5 лет ■ 6 лет (Щ7 лет
б Вилючинская б Моховая б Сероглазка СРМЗ
■ 1 год
_______________Щ2 года
1А 03 года j
_ -йВ_-
,1,1 .il
б Моховая б Сероглгжа СРМЗ
Рис. 3. Особенности распределения средних показателей длины разновозрастных представителей Fucus evanescens в августе (А) и сентябре (Б) 1998 г.
Все представители более старших возрастов (четырех-, пяти- и шестилетние) имели максимальные средние размеры в б. Моховая. В то же время длина семилетних растений уменьшается во всех районах исследований.
Уменьшение длины связано с тем, что именно в период с августа по сентябрь у растений старших возрастных групп по завершении процесса спороно-шения происходит сброс рецептакулов. При этом в верхушечной части фукуса после высева из концептакулов гамет, как показывают наши исследования, резко снижается содержание влаги. Верхушки ветвей, несущие зрелые рецептаку-лы, высыхают и отпадают от слоевищ. Именно это обстоятельство приводит к некоторому снижению средней длины фукусов. Поздней осенью ростовые процессы замедляются, и в зимний период растения находятся в покое.
Исследования, проведенные в 1999 году, позволили проследить особенности роста фукусов у тех же возрастных групп на следующем году вегетации. Сбор количественных проб фукуса был начат с мая (рис. 4). У появившихся проростков средняя длина по-прежнему была максимальной в б. Моховая и составила 0,8 см. В районе СРМЗ и б. Сероглазка — 0,5 и 0,3 см соответственно. Перезимовавшие первогодки, ставшие двухлетними растениями, в б. Моховой
30 25
S 15
s
5
10 ,
i
5 -
i
О — -
и районе СРМЗ увеличили свою длину по сравнению с сентябрем прошлого года (временем последнего сбора проб в 1998 году).
б Моховая б Сероглазка СРМЗ
Рис. 4. Особенности распределения средних показателей длины разновозрастных представителей Fucus evanescens в мае 1999 г.
В б. Сероглазка длина растений в мае 1999 года по сравнению с таковой в сентябре 1998 года уменьшилась. Такая же особенность наблюдалась и у представителей всех возрастных групп в разных районах исследований.
Мы полагаем, что это необычное явление можно объяснить несколькими причинами. Во-первых, воздействием ледового припая. Так как фукус живет в среднем горизонте литорали, то при формировании припая он вмерзает в лед. В период весеннего таяния льда циркулирующая вода расплавляет лед в верхней и нижней части снежно-ледового слоя. Циркуляция воды, ветры обрывают частицы льда, вместе с которыми отрываются верхушки самых высоких растений. А так как фукусы растут не беспорядочно, концентрируются возрастными группами по 1-2 генерации, то в результате разрушения льда под действием гидродинамических условий оборванными оказываются самые высокие растения каждой возрастной группы. Во-вторых, весной средний горизонт литорали большую часть суток находится под водой. Фукус становится одним из наиболее распространенных объектов питания беспозвоночных, наносящих урон его ростовым параметрам. В-третьих, у фукусов, как и у других бурых водорослей, временами наблюдается экскреция органических веществ в окружающую среду (Ерохин, 1981; Камнев, 1989; Клочкова, Трофимова, 2000), что влечет за собой некоторое уменьшение длины.
Результаты исследований динамики изменения длины растений, проведенных в 1999 и 2000 годах, были аналогичны приведенным выше.
ГЛАВА 6. ВОЗРАСТНОЕ И СЕЗОННОЕ НАКОПЛЕНИЕ МАССЫ К ЕУАКЕ.ЪСЕШ В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ
Во всех районах исследования в 1998-2000 годах наряду с показателями длины изучались изменения массы в разные годы и сезоны сбора растений. Динамика изменения массы в июне и июле 1998 года представлена на рис. 5 (А, Б).
8 15 2
10
-гМ d J . J
■ I год В 2 года
□ 3 года
□ 4 года
■ 5 лет
■ 6 лет
■ 7 лет
6 Вилючннская
б Моховая ( б Сероглазка
б Сероглазка
Рис. 5. Особенности распределения средних показателей массы разновозрастных представителей Fucus evanescens в июне (А) и июле (Б) 1998 г.
Из рисунка видно, что масса фукусов в исследуемых районах Авачинской губы различна. Кроме того, при сравнении их массы с массой растений в б. Ви-лючинской заметно, насколько накопление массы у растений из б. Вилючин-ской больше, чем у растений в Авачинской губе.
В августе (рис. 6 А), накопление массы растениями продолжалось. В этот период резкое увеличение массы наблюдалось у растений всех возрастных групп. Увеличение массы объясняется тем, что в августе у фукусов завершается созревание рецептакулов. Рецептакулы увеличиваются в объеме в 10-12 раз,
и общая масса растения увеличивается. Кроме того, растения увеличивают массу за счет верхушечного роста новых ветвей.
В сентябре (рис. 6 Б) у представителей некоторых возрастных генераций наблюдается снижение массы. Так, однолетние фукусы в б. Моховой снижают массу на 3,8%, в районе СРМЗ — на 41,8%, в б. Сероглазка — на 8%. Двухлетние растения в б. Моховой свою массу не изменяют, в районе СРМЗ они становятся тяжелее’на 16,8%, в б. Сероглазка — на 7%. Трехлетние фукусы снижают свою массу во всех районах’исследований. В б. Моховой они становятся легче на 10,3%, в районе СРМЗ — на 5,2%, в б. Сероглазка — на 3,6%. Масса четырехлетних фукусов уменьшаетЬя в б. Моховой на 62,9%, в районе СРМЗ — на 88,4%, в б. Сероглазка — на 66,2%. Пятилетние растения уменьшают массу в б. Моховой на 58,9%, в районе СРМЗ — на 47,3%, в б. Сероглазка — на 53,3%.
6 Вилючияская
б Моховая
б Сероглазка
СРМЗ
б. Меновая
б. Сероглазка
СРМЗ
Рис. 6. Особенности распределения средних показателей массы разновозрастных представителей Fucus evanescens в августе (А) и сентябре (Б) 1998 г.
У растений старших возрастных генераций в сентябре также наблюдается снижение массы, причем происходит этот процесс более интенсивно. Так, масса шестилетних фукусов из б. Моховой снижается на 86,2%, в районе СРМЗ -на 87,5%, в б. Сероглазка — на 88,1%. Семилетние растения уменьшают свою массу в б. Моховой, районе СРМЗ и б. Сероглазка на 83,2%, 85,6% и 86,4% со-
ответственно. Это объясняется теми же причинами: сбросом рецептакулов и частичной экскрецией органических веществ в окружающую среду в указанный период.
Проведенные исследования позволяют судить о том, что в экологически чистых условиях накопление массы слоевищ приходится на период созревания рецептакулов. При этом масса зрелых растений по сравнению с начальной увеличивается наиболее заметно — до 80 раз, наиболее заметно прибавляется масса у пяти- и шестилетних растений. Период созревания рецептакулов в чистой среде продолжается вплоть до первой декады сентября. Здесь растения вступают в размножение на четвертом году жизни. Основной вклад в воспроизводство популяции вносят пяти- и шестилетние представители. Сброс пустых рецептакулов происходит в конце августа — начале сентября. И при этом теряется до 73% общей массы популяции.
В загрязненной среде (бухтах Моховой, Сероглазка и районе СРМЗ) период созревания рецептакулов заканчивается на 15-17 суток раньше, чем в естественных условиях. При этом масса растений увеличивается в 65-70 раз, наиболее значимое прибавление массы наблюдается у шести- и семилетних растений. Здесь растения становятся половозрелыми на третьем году жизни. Сброс пустых рецептакулов заканчивается в последнюю декаду августа. При этом теряется до 75% общей массы популяции. В целом изменение массы у фукусов хорошо сочетается с ходом ростовых процессов у растений.
Динамика изменения массы фукусов в 1999 и 2000 годах была аналогична приведенным выше результатам.
ГЛАВА 7. ВОЗРАСТНАЯ СТРУКТУРА ПОСЕЛЕНИЙ К ЕУАКМСЕМ В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ
Анализ литературных данных показывает, что продолжительность жизни фукусов в пределах ареала значительно варьирует, что определяется сложным сочетанием климатических, гидрологических и гидрохимических факторов. Оптимальные условия их обитания, судя по всему, складываются в нормальных соленых и слабо опресненных водах северных умеренных широт. Несмотря на наличие в настоящее время достаточно обширной литературы по воздействию на виды и популяции фукусов отдельных экологических факторов, определенная точка зрения о влиянии комплекса условий средк на их возрастное развитие пока отсутствует.
Проведенные наблюдения показывают, что в верхних отделах литорали растения более регулярно сталкиваются со стрессовыми ситуациями. В суровые зимы их уничтожает мощный ледовый припай, летом — частые летние дожди или избыточная ультрафиолетовая радиация и сильное пересыхание. Поэтому наиболее полноценную возрастную структуру имеет фукусовое сообщество, развивающееся в нижних горизонтах осушной зоны. Именно на таких участках литорали отбирались все исследованные в ходе работы пробы фукуса.
Определять численность возрастных групп у некоторых видов фукусов достаточно сложно из-за наличия у растений общей подошвы, от которой может отходить несколько десятков вертикальных слоевищ. Иногда под пологом одних видов или на их подошве прорастают зиготы других видов фукуса. На Камчатке, как было указано выше, встречается только один вид фукуса -Р. еуапехсею. Поэтому при проведении исследований подобная проблема не возникала, и подсчет численного состава разновозрастных групп не вызывал особых затруднений.
Первые пробы фукуса в б. Моховой были собраны в первой декаде мая, когда литоральная зона была уже полностью свободна от ледового припая. Результаты обработки этого материала приведены на рисунке 7 А.
I гол 2 года 3 года 4 года 5 лег 6 лет 7 лег
Возрастная группа
I год 2 года 3 года 4 года 5 лет 6 лег 7 лет
Возрастная группа
Рис. 7. Возрастная структура популяции фукуса в б. Моховая в мае (А) и июне (Б) 1999 г.
Из него видно, что в мае доля первогодних растений составила 33,4%. Столь же многочисленными были и двухлетние растения, доля которых в общем объеме пробы составила 32,6%. Присутствие же в популяции трехлетних представителей было почти вдвое меньшим. Их удельный вес составил 19,9%. Численность растений старших возрастов сократилась в 4-30 раз по отношению к численности сеголетков. Так, доля четырех-, пяти-, шести- и семилетних растений составила 9,2; 2,7; 1,2 и 1% соответственно.
В июне (рис. 7 Б) описанные ранее особенности возрастной структуры популяции в целом сохранились. По абсолютной и относительной численности по-прежнему доминировали одно- и двухлетние растения. Их суммарная доля в общем объеме выборки составила 67,7%. По отношению к предыдущему месяцу доля первогодних растений увеличилась на 1,8%, а двухлетних — на 3,4%.
Изменения долевого участия трехлетних фукусов с мая по июнь были направлены в сторону уменьшения и составили 16,1%, в то время как у четырехлетних растений они возросли на 24% по отношению к майской доле вида в общей изученной выборке. Долевое участие пятилетних фукусов, как это видно из рисунка, с мая по июнь сократилось на 26%. Шести- и семилетние представители изучаемого вида не изменили свою численность. Их доля в июньской выборке по-прежнему составила соответственно 1,2 и 1%.
В июле (рис. 8 А) наблюдался небольшой рост числа однолетних представителей.
1 год 2 года 3 года 4 года $ лет
Возрастная группа
6 лет 7 л
1 год 2 гот
3 года 4 года 5 лет
Возрастная группа ^
Рис. 8. Возрастная структура популяции фукуса в б. Моховая в июле (А) и августе (Б) 1999 г.
Их доля в выборке возросла на 3%, кроме того, наблюдалось заметное снижение численности семилетних представителей популяции (60%). Увеличение численности первогодок логичнее всего объяснить поздним развитием перезимовавших проростков, отставших в своем развитии от их основной массы,
начавшей свое развитие еще до формирования зимнего ледового припая. Снижение численности самых старых представителей вида произошло большей частью из-за их гибели, связанной с завершением жизненного цикла растений.
В августе (рис. 8 Б) наблюдалось резкое увеличение численности перво-годних представителей изучаемого вида. Так, доля растений в общем объеме выборки увеличилась на 36,6%. Представители других возрастных групп были ,
не столь многочисленны: доля двухлетних растений составила 31,3%, трехлетних — 12,6%, четырехлетних — 3,3%, пяти-, шести- и семилетних растений — 3,3; 1,1 и 0,5% соответственно. *
В сентябре (рис. 9) вновь наблюдалось незначительное увеличение (на 0,5%) численности первогодних растений, а также увеличение численности шести- и семилетних растений. Последнее изменение объясняется, по-видимому, тем, что к осени образуются очередные дихотомические ветвления слоевища у растений всех возрастных групп, а растения максимальных возрастов, завершающие свой жизненный цикл в начале осени, новых дихотомий не образуют, но к этому времени еще не погибают. Они дополняют численность растений старших возрастных групп.
1 год 2 года 3 года 4 года 5 лет 6 лет 7 лет
Вофастиая груши
Рис. 9. Возрастная структура популяции фукуса в б. Моховая в сентябре 1999 г.
Поэтому в сентябре наблюдался некоторый прирост численности представителей старших возрастов. Так, доля шестилетних растений увеличилась на 54,5%, а семилетних — на 160%.
Сравнительный анализ результатов исследования популяций фукуса из разных районов обитания в конечном итоге позволяет судить об адаптационных стратегиях их развития в условиях неблагоприятной среды и сделать заключение о том, что в чистых условиях обитания критическим периодом выживания фукуса является пятый-шестой годы, в неблагоприятной среде, с хроническим загрязнением — третий-четвертый.
Сильное загрязнение может целиком разрушать нормальную возрастную структуру поселений фукуса, что в свою очередь может указывать на сущест-
вование периодических вспышек повышения загрязнения, связанных с увеличением загрязнения по одному или нескольким ингредиентам.
ГЛАВА 8. ОСОБЕННОСТИ РАСПРЕДЕЛЕНИЯ СУХОГО ВЕЩЕСТВА ПО ТАЛЛОМУ FUCUS EVANESCENS КАК ПОКАЗАТЕЛЬ ЕГО ФИЗИОЛОГИЧЕСКОГО СОСТОЯНИЯ
У берегов юго-восточной Камчатки F. evanescens живет до семи лет, и особенностью его развития является строго упорядоченный рост, в результате которого в течение одного вегетационного сезона у него образуется одно дихотомическое ветвление. Следовательно, все растение фукуса представлено разновозрастными фрагментами, и количество сухого вещества в разных частях слоевища F. evanescens, как показывают наши приведенные ниже исследования, изменяется в зависимости от сезонов года и условий произрастания.
Анализ полученных данных позволяет сделать следующие выводы: наибольшее содержание сухих веществ у растений в возрасте одного-пяти лет из чистых местообитаний наблюдается осенью, у шести- и семилетних растений -весной. Это связано с активным потреблением этих веществ в конце жизненного цикла и их максимальным использованием в ходе формирования репродуктивных органов и созревающих в них половых продуктов и завершением жизненного цикла растения. В антропогенных условиях наименьшая обводненность у растений в возрасте одного-пяти лет наблюдается осенью, а у шести-и семилетних растений — летом, что связано с более короткими сроками их вегетации.
Накопление сухих веществ в разных частях слоевища неравномерно, что свидетельствует об их разной функциональной роли. В чистом районе в весенний период содержание сухих веществ более высокое в нижних частях у пяти-и шестилетних растений, их максимальное содержание наблюдается в верхушечных дихотомиях у шестилетних растений и по всему слоевищу у семилетних растений. Летом содержание сухих веществ увеличивается в нижних частях трех-, четырех- и пятилетних растений и максимальным является у шестилетних растений. Осенью увеличение содержания сухих веществ наблюдается у растений в возрасте от одного до пяти лет во всех частях слоевища. Эти изменения хорошо сочетаются с ходом репродуктивных и ассимиляционных процессов. k
В антропогенной среде содержание сухих веществ на 30% меньше, чем в чистой среде во все сезоны года, что говорит о резком ухудшении у растений физиологического состояния и изменении метаболизма в сторону снижения синтеза запасных веществ и их более активного потребления. В стрессовых условиях содержание сухих веществ достигает уровня, сопоставимого со средним у растений из чистой среды, только у шестилетних растений осенью.
Результаты исследований дают возможность разобраться в ходе сезонных изменений технохимических характеристик качества фукусового сырья и помогают понять особенности биологии развития F. evanescens в каждом из перио-
дов его жизненного цикла. Они позволяют судить о функциональном значении разновозрастных особей в жизни популяции. Кроме того, данные по динамике изменения содержания сухого вещества позволят точнее определять сроки созревания растений и оптимальные сроки их промысла.
ВЫВОДЫ
1. Род Fucus имеет повсеместное распространение в умеренных водах Северного полушария. F. evanescens среди видов рода имеет самый широкий ареал. У юго-восточной Камчатки он демонстрирует высокую экологическую изменчивость, что позволяет изучать его развитие здесь в разных условиях природной и антропогенно измененной среды.
2. Развитие фукуса на первом году жизни завершается практически всегда с образованием дихотомического ветвления. В течение года у F. evanescens образуется одна дихотомия. Возраст фукуса соответствует числу дихотомических ветвлений слоевища. В зависимости от условий обитания продолжительность жизни вида у Камчатки изменяется от 5 до 12 лет, у юго-восточного побережья Камчатки она обычно не превышает 7 лет.
3. Период активного линейного роста у фукуса в ненарушенных условиях обитания охватывает период июнь-август и происходит равномерно. Максимальный прирост длины наблюдается у пятилетних растений. В антропогенных условиях активный рост приходится на период июль-август и происходит скачкообразно. Максимальный прирост длины здесь наблюдается у шестилетних растений, и его величина на 40% меньше, чем у растений из чистых местообитаний.
4. В летний период наибольшее воздействие на рост и развитие фукуса оказывает ритмика приливно-отливных колебаний, при которых представители вида способны сохранять жизнедеятельность в течение 20 часов непрерывного иссушения и 6 часов постоянного солнечного освещения. Смена фенологических фаз развития растений привязана к периодам длительного иссушения и условиям формирования и разрушения ледового припая.
5. Активное накопление массы слоевищ приходится на август-сентябрь и совпадает с периодом созревания рецептакулов. У зрелых растений в чистой среде она по сравнению с ранневесенним периодом увеличивается до 80 раз. Наиболее заметно прибавление массы у растений на пятом и шестом годах жизни. После сброса рецептакулов у фертильных растений наблюдается резкое -, снижение массы, и популяция теряет при этом до 73% от ее максимального значения. В загрязненной среде максимальное накопление массы происходит на
15-17 суток раньше, чем в чистой. При этом масса растений увеличивается !
в 65-70 раз, наибольшее ее прибавление наблюдается у шести- и семилетних растений. Потеря массы популяции после сброса рецептакулов составляет 75%.
6. В ненарушенной природной среде растения вступают в размножение в возрасте 4 лет, созревание рецептакулов продолжается до первой декады сентября. Период высева гамет очень кратковременный, и сброс рецептакулов за-
вершается в сентябре. Основной вклад в воспроизводство популяции вносят растения пятого и шестого годов жизни. В грязной среде растения становятся половозрелыми в возрасте 3 лет. Созревание рецептакулов и их сброс завершаются в последнюю декаду августа. Основной вклад в воспроизводство популяции вносят четырех- и пятилетние растения.
7. В ненарушенной природной среде в разновозрастных популяциях по численности доминируют растения первого и второго годов жизни, в антропогенной среде в период исследований численность растений младших возрастов была заметно меньшей и доминировали четырех-, пяти- и шестилетние растения. Это свидетельствует о том, что при подпороговом уровне загрязнения погибают представители самых младших возрастов. Межгодовые изменения размерно-возрастной структуры популяций определяются как успехом размножения в каждом из годов жизни, так и воздействием внешних факторов. Наиболее резкие изменения она претерпевает ранней весной в период разрушения ледового припая. Основные ее изменения направлены на эллиминацию наиболее крупных растений в каждой возрастной группе в связи с выраженной у фукуса тенденцией группового произрастания одновозрастных растений.
8. Изучение сезонной динамики содержания сухих веществ в отдельных частях растений фукуса из разных условий обитания показывает, что в чистой среде наблюдаются существенные различия ее хода как по возрастам, так и по сезонам года. Сезонный ход изменения содержания сухих веществ по слоевищу у растений разного возраста находится в прямой зависимости от фенологической стадии развития, функциональной роли части слоевища, а также от вклада в воспроизводство популяции той или иной возрастной группы.
9. В загрязненной среде сезонные, возрастные и пространственные (по слоевищу) изменения содержания сухих веществ практически не выражены. Здесь во все сезоны года оно почти на 30% меньше, чем в чистой. Это говорит о резком ухудшении у растений физиологического состояния и изменении метаболизма в сторону снижения синтеза запасных веществ и их более активного потребления. В стрессовых условиях обитания содержание сухих веществ у фукуса заметно повышается только у шестилетних растений шестого года жизни осенью и при этом не достигает среднего уровня содержания сухих веществ у растений из чистой среды.
10. Основные изменения биологии развития фукуса в условиях антропогенного загрязнения сводятся к сокращению сроков вегетации растений, вступлению в размножение молодых представителей популяции, к изменению распределения репродуктивной нагрузки между представителями разных возрастных групп, а также к снижению продукционных показателей. Все эти изменения достаточно различны в местах с разным уровнем загрязнения, носят адаптивный характер и направлены на сохранение вида в условиях стрессовых нагрузок и могут использоваться для мониторинга состояния прибрежных вод.
11. Данные изучения биологии развития фукуса показывают, что в ходе его промысла целесообразно выборочное изъятие растений 5-6 годов жизни в период июля-августа, когда у фукуса наблюдается максимальное содержание
сухих веществ и максимальная биомасса. Период эксплуатации одной популяции не должен превышать 3-х лет и для обеспечения нормального воспроизводства фукуса объем изъятия не должен превышать 30% от общего количества растений, произрастающих в районе сбора.
СПИСОК РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Чмыхалова В.Б. Развитие бурой водоросли Fucus.evanescens в прикамчатских водах // Тез. докл. науч.-технич. симпозиума «Современные средства воспроизводства и использования водных биоресурсов». — С.-Пб., 2002. -Т.1. — С. 68-71.
2. Трофимова Т.Н., Чмыхалова В.Б. Развитие массовых видов бурых водорослей камчатского шельфа Laminaria bongardiana и Fucus evanescens в разных экологических условиях в летний период // Тез. докл. междунар. конф. «Биологические основы устойчивого развития прибрежных морских экосистем». — Мурманск, 2001. — С. 238-240.
3. Березовская В.А., Чмыхалова В.Б. Видовое разнообразие сообщества Fucus evaanescens, его продукционные характеристики и размерно-возрастная структура поселений в Авачинской губе // Матер. 2-й науч. конф. «Сохранение биоразнообразия Камчатки и прилегающих морей». — Петропавловск-Камчатский, 2001. — С. 26-28.
4. Чмыхалова В.Б. Результаты изучения размерно-возрастной структуры популяции Fucus evanescens (Ag.) на острове Парамушир // Матер. 3-й науч. конф. «Сохранение биоразнообразия Камчатки и прилегающих морей». — Петропавловск-Камчатский, 2002. — С. 298-299.
5. Клочкова Н.Г., Чмыхалова В.Б. Продукционные характеристики Fucus evanescens и размерно-возрастная структура его поселений в Авачияской губе в летний период // Матер, науч.-технич. конф. «Ресурсы и средства рациональной эксплуатации прибрежных акваторий Камчатки». — Петропавловск-Камчатский, 2003. — С. 99-102.
6. Чмыхалова В.Б., Королева Т.Н. Некоторые особенности вегетации бурых водорослей Laminaria bongardiana и Fucus evanescens в разных районах Авачинского залива // Матер, науч.-технич. конф. «Ресурсы и средства рациональной эксплуатации прибрежных акваторий Камчатки». — Петропавловск-Камчатский, 2003. — С. 40-44.
7. Клочкова Н.Г., Чмыхалова В.Б., Королева Т.Н. Биология, экология и распространение рода Fucus и вида Fucus evanescens Ag. // Матер. I и II сессии Русского бот. общества. — Петропавловск-Камчатский, 2004. — С. 68-87.
8. Чмыхалова В.Б. Использование бурой водоросли Fucus evanescens для оценки экологического состояния прибрежных вод юго-восточной Камчатки // Известия высших учебных заведений. Северо-Кавказский регион. Прил. № 4. — Новочеркасск, 2004. — С. 67-70.
9. Чмыхалова В.Б., Королева Т.Н. Динамика содержания сухого вещества у разновозрастных представителей камчатской бурой водоросли Fucus evanes-
cens в Авачинском заливе // Матер. 5-й науч. конф. «Сохранение биоразнообразия Камчатки и прилегающих морей». — Петропавловск-Камчатский, 2004. -С. 34-36.
10. Чмыхалова В.Б., Королева Т.Н. Влияние среды обитания на развитие бурой водоросли Fucus evanescens Ag. // Матер, региональной науч.-практич. конференции. — Петропавловск-Камчатский, 2004. — С. 56-58.
11. Чмыхалова В.Б., Королева Т.Н. Особенности сезонного распределения сухих веществ в различных частях растений Fucus evanescens Ag. // Матер, науч.-практич. конференции проф.-препод. состава и аспирантов 20-27 апреля 2004 г. — Петропавловск-Камчатский, 2005. — С. 94-103.
I
I
Чмыхалова Виктория Борисовна
РАЗВИТИЕ БУРОЙ ВОДОРОСЛИ FUCUS EVANESCENS Ag. В ПРИКАМЧАТСКИХ ВОДАХ
Автореферат диссертации на соискание ученой степени кандидата биологических наук
Редактор И.В. Скрыпкина Технический редактор Е.Е. Бабух Набор текста, верстка В.Б. Чмыхалоэа Оригинал-макет Е.Е. Бабух
Подписано в печать 29.09.2005 г. ‘ Формат 61 *86/16. Печать офсетная. Гарнитура Times New Roman Авт. л. 1,05. Уч.-изд. л. 1,27. Усл. печ. л. 1,69 Тираж 110 экз. Заказ № 483
Редакционно-издательский отдел Камчатского государственного технического университета
Отпечатано полиграфическим участком РИО КамчатГТУ 683003, г. Петропавловск-Камчатский, ул. Ключевская, 35
»18 5 22
РНБ Русский фонд
2006-4 19623
Библиография Диссертация по биологии, кандидата биологических наук, Чмыхалова, Виктория Борисовна, Петропавловск-Камчатский
1. Барашков Г.К. Сравнительная биохимия водорослей. М.: Пищевая промышленность, 1972. -336 с.
2. Бек Т.А., Потапова Л.И. Разложение макрофитов в литоральной зоне Белого моря // Экология, 1986. № 1. С. 79-82.
3. Березовская В.А. Гидрохимический режим Авачинской губы // Авто-реф. дис. канд. геогр. наук. Ростов-на-Дону, 1988. — 25 с.
4. Березовская В.А. Авачинская губа. Гидрохимичекий режим, антропогенное воздействие. Петропавловск-Камчатский: КГ АРФ, 1999. — 156 с.
5. Березовская В.А. Накопление свинца бурой водорослью Laminaria bongardiana в Авачинской губе // Матер, конф. проф.-препод. состава и аспирантов 1999-2000 гг. Тр. КамчатГТУ. Петропавловск-Камчатский: КамчатГТУ, 2001. ВыпЛЗ.С. 7-11.
6. Березовская В.А. Макрофитобентос как показатель состояния среды в прибрежных водах Камчатки // Автореф. дис. докт. геогр. наук. Владивосток, 2002. — 49 с.
7. Березовская В.А., Клочкова Н.Г. Содержание кислорода, биогенных и загрязняющих веществ в воде Авачинской губы // Сб. науч. статей по экологии и охране окружающей среды Авачинской бухты. Петропавловск-Камчатский: Изд-во Госкамчатэкология, 1998. С. 19-26.
8. Блинова Е.И. Вертикальное распределение и количественный учет мак-рофитов Айновских островов (Баренцево море) // Тр. Мурманского морского биологического института. Л.: Наука, 1965а. Вып. 8/12. С. 41-55.
9. Блинова Е.И. Распределение и количественные показатели промысловых водорослей сублиторали Мурмана (Баренцево море) // Растительные ресурсы, 19656. Т. 1, Вып. 4. С. 583-589.
10. Блинова Е.И. Морские водоросли северо-восточной части Охотского моря // Новости сист. низш. раст. Л., 1968а. С. 33-38.
11. Блинова Е.И. Видовой состав и вертикальное распределение морских водорослей в Пенжинской губе (Охотское море) // Океанология, 19686. Т. 8, Вып. 2. С. 279-286.
12. Блинова Е.И. Морские водоросли северо-восточной части Охотского моря / Морские подводные исследования. М.: Наука, 1969. С. 114-120.
13. Блинова Е.И., Возжинская В.Б. Морские макрофиты и растительные ресурсы океана // Основы биол. продуктивности океана и ее использование. -М.: Наука, 1971.
14. Богданов К.Т., Мороз В.В. Структура, динамика и гидролого-акустические характеристики вод проливов Курильской гряды. Владивосток: Дальнаука, 2000. — 152 с.
15. Вербина Н.М. Гидромикробиология. М.: Пищевая промышленность, 1980.-283с.
16. Витюк Д.М. Взвешенное вещество и его биогенные компоненты. -Киев: Наук, думка, 1983. 210 с.
17. Виноградова К.Л. Видовой состав водорослей литорали о-ва Куба // Новости сист. низш. раст. Л.: Наука, 1975. С. 94-101.
18. Виноградова К.Л. Водоросли юго-западного побережья Берингова моря // Новости сист. низш. раст. Л.: Наука, 1978. С. 3-11.
19. Виноградова К.Л., Клочкова Н.Г., Перестенко Л.П. Список водорослей литорали восточной Камчатки и западной части Берингова моря // Литораль Берингова моря и юго-восточной Камчатки. М.: Наука, 1978. С. 150-155.
20. Возжинская В.Б. Распределение водорослей на литорали бухты Грос-севичи (Японское море) // Бот. журнал. М.: Наука, 1964. С. 712-714.
21. Возжинская В.Б Морские водоросли западного побережья Камчатки // Новости сист. низш. раст. Л.: Наука, 1965. С. 73-78.
22. Возжинская В.Б., Селицкая (Вишневская) Н.М. Видовой состав, распределение и запасы фукоидов в Охотском море // Тр. Института океанологии, 1970. Т. 88. С. 281-288.
23. Возжинская В.Б. К продуктивности макрофитобентоса Охотского моря // Всесоюзное совещ. по морской альгологии-макробентосу: Тез. докл. М., 1974. С. 31-33.
24. Возжинская В.Б. Сезонные изменения макрофитобентоса Кандалакшского залива Белого моря. М., 1980. С. 85-93.
25. Возжинская В.Б. Донные макрофиты Белого моря. М.: Наука, 1986. -190 с.
26. Возжинская В.Б. Макрофитобентос высокобореальных (холодновод-но-умеренных) морей России // Автореф. дис. докт. биол. наук. М., 1996 -45 с.
27. Возжинская В.Б., Цапко A.C., Блинова Е.И., Калугина A.A., Петров Ю.Е. Промысловые водоросли СССР. Справочник. М. : Пищ. пром-сть, 1971 — 270 с.
28. Возжинская, Ярцева, Максимова Особенности суммарного фотосинтеза некоторых фукоидов Белого и Японского морей // Экология фауны и флоры прибрежных зон океана. М.: Институт океанологии им. П.П. Ширшова АН СССР, 1985. С. 46-52.
29. ГОСТ 7636-85. Рыба, морские млекопитающие, морские беспозвоночные и продукты их переработки. Методы анализа // Государственные стандарты М.: изд-во стандартов, 1998. С. 36 — 121.
30. Гринталь А.Р. Состав и распределение сообществ водорослей на литорали губ Ярнышной и Подпахты (Восточный Мурман) // Распределение и состав промысловых водорослей Баренцева моря. М-Л.: Наука, 1965. С. 23-40.
31. Гурьянова Е.Ф. Командорские острова и их морская прибрежная фауна и флора. Природа, 1935. № 11. С. 64-72.
32. Гусарова И.С. Макрофиты сублиторальной зоны островов Итуруп, Уруп, Симушир (Большая Курильская гряда) // Новости сист. низш. раст. Л., 1975. С. 111-118.
33. Гусарова И.С. Видовой состав макрофитов б. Рудная (Японское море) // Новости сист. низш. раст. Л.: Наука, 1982. С. 3-16.
34. Гусарова И.С. Видовой состав макрофитов лагуны Семячик (Юго-восточная Камчатка) // Новости сист. низш. раст. Л.: Наука, 1987. С. 22-25.
35. Евсеева Н.В. Состояние зарослей и ресурсы промысловых водорослей в прибрежье островов Малой Курильской гряды // Растительные ресурсы, 1997. Вып. 4.-С. 98-104.
36. Ерохин В.Е., Карнаухов В.Н. Состояние энергетического аппарата макрофитов в норме и при фенольной интоксикации // Экология моря. —1981. — Вып. 6. С. 61-66.
37. Ефименко А.К. Предисловие // Сб. науч. ст. по экологии и охране окружающей среды Авачинской бухты. -Петропавловск-Камчатский-Токио, 1998. С. 3-5.
38. Зенин A.A., Березовская В.А. Антропогенное воздействие на загрязненность Авачинской губы и ее притоков // Тез. докл. 28 Всесоюз. гидрохим. совещ. Ростов-на-Дону, 1984. С. 18.
39. Зинова А.Д. Список морских водорослей южного Сахалина и южных островов Курильской гряды // Исслед. дальневост. морей СССР. 1959. Вып. 6. С. 146-161.
40. Зинова А.Д. К вопросу о фитогеографическом районировании прибрежной полосы Мирового океана // Тез. докл. конф. по совместным исслед. флоры и фауны. JL, 1962. С. 1-9.
41. Зинова А.Д. Новые данные о морских водорослях из чаунской губы (Восточно-Сибирское море) // Новости сист. низш. раст. Л.: Наука, 1970. С. 102-107.
42. Зинова Е.С. Водоросли Охотского моря с побережий Большого Шан-тарского острова // Тр. Ленингр. о-ва естествоисп. 1930. Т. 60, Вып. 3. С. 81125.
43. Зинова Е.С. Морские водоросли юго-восточной Камчатки // Тр. Ботан. ин-та АН СССР, 1954. Сер. 2, Вып. 9. С. 365-400.
44. Иванюшина Е.А. Изменения в бентосе Авачинской губы под влиянием антропогенного загрязнения // Сб. науч. статей по экологии и охране окружающей среды Авачинской бухты. Петропавловск-Камчатский: Изд-во Гос-камчатэкология, 1998. С. 47-51.
45. Камнев А.Н. Структура и функции бурых водорослей. М.: Изд-во МГУ, 1989.- 198 с.
46. Кацыка А.П. Климат // Сб. статей и очерков по географии. Петропавловск-Камчатский, 1966. С. 60-66.
47. Кизеветтер И.В., Грюнер B.C., Евтушенко В.А. Переработка морских водорослей и других промысловых водных растений. -М.: Пищевая промышленность, 1967. 416 с.
48. Клочкова Н.Г. Видовой состав водорослей литорали и сублиторали Камчатского залива // Новости сист. низш. раст. Л., 1976. Т. 13. С. 20-24.
49. Клочкова Н.Г. Аннотированная библиография по морским водорослям-макрофитам Татарского пролива (Японское море). (Первая ревизия флоры). Владивосток: Дальнаука, 1994. — 108 с.
50. Клочкова Н.Г. Флора водорослей-макрофитов Татарского пролива и особенности ее формирования. Владивосток: Дальнаука, 1996. 288 с.
51. Клочкова Н.Г. Водоросли-макрофиты дальневосточных морей России // Автореф. дис. докт. биол. наук. Владивосток, 1998. — 45 с.
52. Клочкова Н.Г., Березовская В.А. Антропогенная трансформация морской бентосной растительности в Авачинской губе (юго-восточная Камчатка) // Тез. докл. Всерос. съезда «Растения в умеренном климате». Владивосток: Дальнаука, 1998. С. 129-131.
53. Клочкова Н.Г., Березовская В.А. Макрофитобентос Авачинской губы и его антропогенная деструкция. Владивосток: Дальнаука, 2001 — 208 с.
54. Клочкова Н.Г., Трофимова Т.Н. Воспроизводство массовых ламинариевых водорослей в водах Камчатки // Годовой отчет о НИР. Петропавловск-Камчатский: КамчатНИРО, 2000. — 37 с. (рукопись)
55. Ковальчук Н.А. Видовой состав и биомасса макрофитов цистозиры из акваторий нескольких Крымских пляжей. Альгология, 1992. Т. 2. С. 48-52.
56. Кондратюк В.И. Климат Камчатки. М.: Гидрометеоиздат, 1974.202 с.
57. Константинов А.С. Общая гидробиология. М.:Высшая школа, 1986.472 с.
58. Копылов Б.И., Павлова В.П. Экология Авачинской губы: источники загрязнения, проблемы, решения, перспективы // Сб. науч. статей по экологии и охране окружающей среды Авачинской бухты. Петропавловск-Камчатский: Изд-во Госкамчатэкология, 1998. С. 11-18.
59. Кузнецов В.В. Влияние зимнего ледяного припая на морфологию побережья фауны и флоры литорали Белого моря // Работы Мор. биол. станции, 1948. Вып. 1. С. 44-57.
60. Кузнецов В.В. Белое море и биологические особенности его флоры и фауны. М.; Л.: Изд-во АН СССР, 1960. 322с.
61. Кузнецов В.В. Биологические особенности основных представителей беломорской флоры и условия их промышленного использования // Тр. Всесо-юз. Совещ. Работников водорослевой промышленности. Архангельск, 1962. С. 131-140.
62. Кудряшов В.А., Тараканова Т.Ф., Иванова М.Б. О фауне и флоре оуш-ной зоне Шантарских островов // Прибрежные сообщества дальневост. морей. — ДВНЦ АН СССР, 1976. Сб. 6. С. 22-63.
63. Кусакин О.Г. Некоторые закономерности распределения фауны и флоры в осушной зоне Южных Курильских о-вов // Исслед. дальневост. морей СССР, 1961. Вып. 7. С. 312-343.
64. Кусакин О.Г. Состав и распределение макробентоса в осушной зоне о-ва Симушир Курильской гряды // Прибрежные сообщества дальневост. морей. ДВНЦ АН СССР, 1976. Сб. 6. С. 5-И.
65. Кусакин О.Г., Иванова М.Б. Беринговоморская литораль Чукотки // Литораль Берингова моря и юго-восточной Камчатки. М.: Наука, 1978. С. 10-40.
66. Кусакин О.Г., Иванова М.Б. Макрофитобентос литоральных сообществ о-ва Медный (Командорские острова) // Биол. моря, 1995. Т. 21, № 2. С. 99-107.
67. Кукк Х.А. Новые водоросли острова Ытыгран (Берингово море) и Ко-лючинской губы (Охотское море) // Новости сист. низш. раст. Л.: Наука, 1984. С. 23-25.
68. Ле Нгуен Хиеу Морские водоросли северного Вьетнама // Новости сист. низш. раст. Л.: Наука, 1970. С. 71-81.
69. Макаров М.В. Влияние ультрафиолетовой радиации на рост и размножение доминантных видов водорослей-макрофитов Баренцева моря // Ав-тореф. дис. канд. биол. наук. С.-Пб., 1999. — 22 с.
70. Макиенко В.Ф. Водоросли-макрофиты залива Восток (Японское море)
71. Биология моря, № 2, 1975. С. 45-47.
72. Максимова Н.Я. Моря, омывающие Камчатку // Сб. статей и очерков по географии. Петропавловск-Камчатский, 1966. С. 17-24.
73. Максимова О.В. Некоторые сезонные особенности развития и определения возраста беломорских фукоидов // Донная флора и продукция краевых морей СССР. М.: Наука, 1980. С. 73-78.
74. Малиновская Т.М. Содержание металлов в разновозрастных талломах бурой водоросли Fucus evanescens Курильских островов // Биология моря, 1998. -Т. 24, №6.-С. 397-399.
75. Мейер К.И. Материалы по флоре водорослей Белого моря // тр. ВНИ-РО.-М., 1938, т. 7. С. 5-28.
76. Мелекесцев И.В. Рельеф // Сб. статей и очерков по географии. Петропавловск-Камчатский, 1966. С. 25-31.
77. Мессинева Е.М. Возрастная морфофизиология промысловых представителей семейства Cystoseiraceae морей России // Автореферат на соискание ученой степени кандидата биологических наук. -М., 2003,22с.
78. Михайлова Н.Ф. Распределение высших водорослей вдоль берегов о-ва Шикотан // Ботан. журн. 1959. Т. 44, N 3. С. 379-386.
79. Михайлова Т.А. Развитие ламинариевых фитоценозов на внесенном каменистом субстрате в Белом море // Автореф. дис. канд. биол. наук. С-Пб, 2000. — 20 с.
80. Перестенко Л.П. Распределение водорослей на литорали губ Плохие и Большие Чевры (Восточный Мурман) // Распределение и состав промысловых водорослей Баренцева моря. М.-Л.: Наука, 1965. С. 13-22.
81. Перестенко Л.П. Водоросли залива Петра Великого. Л.: Наука, 1980.232 с.
82. Перестенко Л.П. Фитоценозв сублиторали юго-западного побережья Охотского моря и Шантарских островов // Бот. журн., 1996. Т. 81. №7. С. 41100.
83. Петров Ю.Е. Fucus distichus L. Emend Powell и F. evanescens Ag. // Новости сист. низш. раст. Л.: Наука, 1965. С. 64—70.
84. Петров Ю.Е. Ламинариевые и фукусовые водоросли морей СССР // Автореф. дис. докт. биол. наук. Л., 1975. — 53 с.
85. Петров Ю.Е. Отдел Бурые водоросли (Phaeophyta) // Жизнь растений. Т. 3. Водоросли, лишайники. -М.: Просвещение, 1977. С. 143-191.
86. Постельс А., Рупрехт Ф.И. Изображения и описания морских растений, собранных в северном Тихом океане у берегов Росийских владений в Азии и Америке. СПб., 1840 — 22 с.
87. Силкин В.А., Хайлов K.M. Биоэкологические механизмы управления в аквакультуре. Л.: Наука, 1988 — 230 с.
88. Спасский H.H. Литораль юго-восточного побережья Камчатки // Ис-след. дальневост. морей СССР, 1961. Вып. 7. С. 261-311.
89. Суховеева М.В. Распределение водорослей вдоль берегов Приморья // Известия ТИНРО. -В.: Книжное издательство, 1967. С. 255-260.
90. Тараканова Т.Ф. Количественное распределение макробентоса на литорали острова Беринга // Литораль Берингова моря и юго-восточной Камчатки. М.: Наука, 1978. С. 63-77.
91. Тиховская З.П. Видовой состав морских водорослей в районе Мурманской биологической станции // Тр. Мурманской. Биол. станции, 1948. С. 189-191.
92. Толстикова Н.Е. Распределение водорослей и характеристика состояния запасов промысловых водорослей в Анадырском заливе // Научный отчет. -М.: ВНИРО, 1973-92 с.
93. Толстикова Н.Е. Новые данные по экологии сублиторальных макро-фитов в Анадырском заливе Берингова моря // Новости сист. низш. раст. Л., 1974. Т. 11. С. 147-152.
94. Усов А.И., Клочкова Н.Г. Бурые водоросли Камчатки как источник маннита//Биоорганическая химия, 1994. Т. 20, № 11. С. 1236-1241.
95. Ушаков П.В. Фауна Охотского моря и условия ее существования. -АН СССР, 1953. С. 317-326.
96. Хайлов K.M. Прижизненное выделение органических веществ морскими макрофитами и экологические условия прибрежной зоны // Тр. Мурм. морского биол. института, 1964. Вып. 5(9). С. 49-56.
97. Хайлов K.M., Парчевский В.П. Иерархическая регуляция структуры и функций морских растений. Киев: Наукова думка, 1983. — 254 с.
98. Христофорова Н.К. Использование бурых водорослей фукусов для индикации состояния прибрежных морских вод // В кн.: Сихотэ-Алиньский биосферный район: принципы и методы биологического мониторинга. Вла1 дивосток: ДВНЦ АН СССР, 1981. С. 118-127.
99. Христофорова Н.К. Биоиндикация и мониторинг загрязнения морских вод тяжелыми металлами. Л.: Наука, Ленинградское отделение, 1989. -192 с.
100. Христофорова Н.К., Малиновская Т.М. Содержание металлов в фукусах бухты Кратерной (Курильские острова) в связи с условиями существования //Биол. моря, 1995. Т. 21. № 1. С. 77-82.
101. Чмыхалова В.Б. Результаты изучения размерно-возрастной структуры популяции F. evanescens (Ag.) на острове Парамушир // Мат. 3 науч. конф. «Сохранение биоразнообразия Камчатки и прилегающих морей». Петропавловск-Камчатский, 2002. — С.298-299.т
102. Шошина Е.В. Дополнение к флоре водорослей Белого моря // Новости сист. низш. раст. Л., 1979. С. 28-33.
103. Щапова Т.Ф. Литоральная флора материкового побережья Японского моря // Тр. Института океанологии, 1957. Т. 23. С. 21-66.
104. Щапова Т.Ф., Возжинская В.Б. Водоросли литорали западного побережья Сахалина // Тр. Института океанологии, 1960. Т. 34. С. 123-146.
105. Agardh С.A. Species algarum rite cognitae cum synonymis differentiis specificis et descriptionibus succinetis // Lund. 1820. vol. 1, part 1. 268 p.
106. Bolton J.J., Liming K. Optimal growth and maximal survival temperaturesof Atlantic Laminaria species (Phaeophyta) in culture // Mar Biol., 1982. Vol. 66. P.89-94.
107. Cheney D.R. The determining effects of snail herbivore density of intertidal algal recruitment and composition // Abstr. 1st. Int. Phycol. Congr. Newfoundland, 1982. P. 48.
108. Clayton M.N, Evolution of the Phaeophyta with particular reference to the Fucales I I Progress in Phycological Research. Biopress Ltd., 1984. Vol. 3. — P. 11-46.
109. Fritsch F.E. The structure and reproduction of the algae // Cambridge, 1945.-936 p.
110. Gardner N.L. The genus Fucus on the pacific coast of North America // Univ. Calif Publ. Botan. V. 10,1922, N 1. 180 p.• > > V
111. Graham L.E., Wilcor l.W. Algae. Prentice hall, upper, saddle river, NJ, 07485. USA: University of Wisconsin, 2000. — 640 p.
112. Gerard V.A., Du Bois K.R. Temperature adaptation in a southern boundary of the kelp Laminaria saccharina // Mar Biol., 1988. Vol. 97. P. 575-580.
113. Hansen G.I. A morphological study of Fibriofolim, a new genus in the Cystocloniaceae (Gigartinales, Rhodophyta) II. Phycol., 1980. № 16. P. 207-217.
114. Hansen G.I. A revised cheklist and preliminary assessment of the macro-benthic marine algae and seagrasses of Oregon // in book: Conservation and Management of Native Flora and fungi. Nativ Plant Society of Oregon, Corvallis. 1997. P. 175-200.
115. Jensen A. Morphological studies in Cystoseiraceae and Sargassaceae (Phaeophyceae) with special reference to apical organization // Univ. Calif. Publ. Bot., 1974. №68. P. 1-61.
116. Keser M., Vadas R., Larson B. Regrowth of Ascophyllum nodosum and Fucus vesiculosus under variosharvesting regimes in marine. USA // Bot. Mar.-1981-V. 24.-P. 29-38.
117. Klochkova N.G. An annotated bibliography of marine macroalgae of the northwest coast of the Bering Sea and southeast Kamchatka. First Revision of Flora // Algae. Formerly the Korean Journal of Phycol., 1998. Vol. 9, № 5 — 90 p.
118. Knight M., ParkeV. A biological study of Fucus vesiculosus and Fucus serratus // J. Mar. Biol. Ass. 1950. -V. 29. №2, -P. 439-514.
119. Levring T. Potential yields of Marine algae with emphasis on European Species // In book: The Marine plant Biomass of the Pacific Northwest Coast. Ed. R. W. Krauss. Oregon State University Press. 1977. p. 251- 270. *
120. Lindstrom S.C. An annotated bibliography of the benthic marine algae of Alaska // Alaska department of Fish and Game, 1977. 172 p.
121. Linneaeus C. Species plantarum, exibentes plantas rite cognitas, ad genera relatas, cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus secundum systema sexuale digestas//Stockholm. 1753. Ed. 1.378 p.
122. Los Petes P.I. Aanmerkingen over de focus soorten van de Belgishe kust // Dumortiera, 1982, N 23. P. 9-16.
123. Moss B. Growth and regeneration of Fucus vesiculosus in culture // Brit. Phycol. Bull. 1964. Vol. 2. P. 5-6.
124. Muller-stoll W.r., Kunzenbach R. Uber die standortbedingten wachformen von Fucus vesiculosus in den Gewässern der Insel Hiddensee // Arch. Fur. Protis-tenkunde. -1956. B. 101. Hs. P. 289-334.
125. Munda S. On the chemical composition, distribution and ecology of some N common benthic marine algae from Iceland // Botanica Marina, 1972, V.XXV, 31, 24-41 p.
126. O’Klair R.M., Lindstrom, S.C. North Pacific Seaweeds. Auke Bay Alaska: Plant Press, 2001. 159 p.
127. Parker J. Seasonal changes in coldhardiness of Fucus vesiculosus II Biol. Bull. 1960. Vol. 119, № 2, P. 474-478.
128. Printz h. Algenvegetation des Trondjemsfjordes // Skr. Norske Vidensk. Acad. Oslo. J. Mar. Nat. KL. — 1926. -№ 5. — 273 S.
129. Phinney H.K. Macrophytic Marine algae of Oregon // In book: The Marine plant Biomass of the Pacific Northwest Coast. Ed. R. W. Krauss. Oregon State University Press. 1977. p.93-115.
130. Powell H.T. Speciation in the genus Fucus L.35.
133. Schonbeck M., Norton T.A. Factors controlling the upper limit of fucoid algae on the shore. // J. Exp. Mar. Biol. Ecol., 1978. P. 303-330.
134. Sj0tun K., Fredriksen S. Growth allocation in Laminaria hyperborea (Laminariales, Phaeophyta) in relation to age and wave exposure // Marine Ecology Progress series, 1995. Vol. 126. P. 213-222.
135. South G.R., Wittick A. Introduction to Phycology // Blackwell Scientific Publication. Oxford London Edinburgh, 1987. P. 74-88.
136. Stomps T.J. Stüdes topographiques sur la variabilité des Fucus vesiculosus L., plalycarpus Thür, et seranoides L // Ree. Inst. Bot. 1911. «Leo Errera», Brüx., 8.-P. 325-377.
137. Tan I.H. Druehl L.D. A ribosomal DNA phylogeny supports the close evolutionaryrelationships among the Sporochnales, Desmsrestiales, and Laminariales (Phaeophyceae) // J. Phycol. 1996. — 32, N 1. — P. 112-118.
138. Tseng C.K. Common seaweeds of China. China, Beijing: Science press, 1983.-316 p.
139.52.»
Водоросли морские «Фукус» сушеные пищевые молотые Водорослевая соль 100г
{{/if}}
{{if typeof(brands) != ‘undefined’}}
${brands.label} — ${priceFormatSpaced(brands.count)}
{{/if}}
{{if typeof(products) != ‘undefined’}}
${products.label} — ${priceFormatSpaced(products.count)}
{{/if}}
{{else}}
Ничего не найдено
{{/if}}
Фильтровать продукты по категориям
СМОТРЕТЬ выбранные категории
Игнор
Добавить в игнор
Удалить из игнора
от 130 00
Добавить в корзину
Убрать из корзины
Всего фото — 2
{{=it.photoTitle || ‘отзыв’}}{{? it.dateCreated }} от {{=it.dateCreated}}{{?}}
{{? it.totalCount > 1 }}
{{=it.index}} / {{=it.totalCount}}
{{?}}
ООО "Гарнец".
Водоросли морские "Фукус" сушеные пищевые молотые Водорослевая соль 100г.Природный источник йода.
Состав: водоросли …
Ваше местоположение
Закрыть
ЕГО ПРОИСХОЖДЕНИЕ, ЭКОЛОГИЯ И ТАКСОНОМИЧЕСКИЕ ПОСЛЕДСТВИЯ ДЛЯ JSTOR
Абстрактный
Морфология и среда обитания карликовых мохоподобных или мускоидных фукоидных бурых водорослей были изучены в солончаке Брейв-Боут-Харбор в Йорк-Киттери, штат Мэн, США, с использованием трансектантных исследований и экспериментов по пересадке растений. Растение, которому не хватает фиксатора, образует плотный заросший дерн среди редких популяций Spartina patens на высоких болотах, особенно на хорошо дренированных песчаных отложениях у устья гавани.Дихотомически разветвленные листья растения были меньше, чем те, о которых ранее сообщалось для мускоидоподобных популяций из Европы (в среднем = 13,2 мм в длину, 1,1 мм в ширину и 0,1 г влажно-высушенного веса), в то время как у них было такое же преобладание краевых ямок для волос или криптостом . Пересадка in situ Fucus spiralis с нижних на верхние болота привела к усилению фрагментации, задержке роста, пролиферации и снижению воспроизводства. Взаимная трансплантация мускоидоподобного фукуса с верхних на нижние возвышения вызвала увеличение длины и разрастания вайя.На основании подробных трансплантационных и морфологических исследований мы пришли к выводу, что мускоидоподобные растения фукуса из Brave Boat Harbor представляют собой фенотипический вариант F. spiralis, вызванный отслоением, обширным разрастанием и последующей дегенерацией отделившихся фрагментов. Карликовая морфология растения в первую очередь связана с рядом уникальных условий окружающей среды (высыхание и низкий уровень питательных веществ), а также с типом доступного родительского материала. Таким образом, карликовый мускоидоподобный фукус в Европе и на некоторых участках Северо-Западной Атлантики может происходить от F.vesiculosus, а в гавани Брейв-Боут родительский материал — F. spiralis. Аналогичная картина также очевидна между Ascophyllum nodosum и его обособленным ecad scorpioides, с присутствием карликовых экземпляров и появлением заметного морфологического континуума между двумя растениями.
Информация о журнале
Rhodora — рецензируемый ботанический журнал, посвященный главным образом флоре Северной Америки. Он постоянно публикуется с 1899 года.Этот рецензируемый ежеквартальный отчет состоит из 400-500 страниц в год. Будут рассмотрены монографии или научные статьи, касающиеся систематики, флористики, экологии, физиологии, палеоботаники или природоохранной биологии флоры Северной Америки или связанных с флористикой областей.
Информация об издателе
Родора издается ежеквартально Ботаническим клубом Новой Англии (NEBC) и публикуется совместно с Allen Press, Inc. NEBC, основанная в 1895 году, является некоммерческой организацией, которая способствует изучению растений Северной Америки, особенно флоры Новой Англии. Англия и прилегающие районы.Клуб проводит ежемесячные встречи в течение учебного года (обычно в Гарвардском университете, Кембридж, Массачусетс), поддерживает гербарий из более чем 253000 листов, имеет небольшую библиотеку и ежегодно присуждает студентам научную премию и премию Фернальда за публикации.
Сезонные колебания плодовитости Fucus vesiculosus при закислении океана и потеплении в западной части Балтийского моря
Потепление и закисление океана может существенно повлиять на воспроизводство ключевых видов, таких как Fucus vesiculosus (Phaeophyceae).В четырех последовательных экспериментах с бентосным мезокосмом мы сравнили репродуктивную биологию и количественно оценили временное развитие плодородия Балтийского моря Fucus под единичным и комбинированным воздействием повышенной температуры морской воды и pCO 2 (1100 частей на миллион). В дополнительном эксперименте мы исследовали влияние температуры (0–25 ° C) на созревание сосудов F. vesiculosus в Северном море. Выраженный сезонный репродуктивный цикл F. vesiculosus проявился в течение 1 года.На первое появление цветоложов на вегетативных верхушках и дальнейшее развитие незрелых цветоносов у F. vesiculosus осенью не повлияло потепление или повышение pCO 2 . Зимой повышенное значение pCO 2 как при температуре окружающей среды, так и при повышенных температурах значительно увеличило долю зрелых сосудов. Весной потепление и, в меньшей степени, повышение pCO 2 ускоряли созревание цветоносов и увеличивали высвобождение гамет на срок до 2 недель.Аналогичным образом, в лаборатории созревание и высвобождение гамет ускорялись при 15–25 ° C по сравнению с более низкими температурами. Таким образом, повышенный pCO 2 и / или потепление не влияют на внешний вид цветоносов осенью, но ускоряют процесс созревания весной, что приводит к более раннему высвобождению гамет. Температура и, в гораздо меньшей степени, pCO 2 влияют на временное развитие фертильности Fucus . Таким образом, повышение температуры в основном сместит или нарушит фенологию F.vesiculosus весной и летом, что может изменить и / или затруднить его экологические функции в мелководных прибрежных экосистемах Балтийского моря.
Ссылки
Åberg, P. and H. Pavia. 1997. Временные и многомасштабные пространственные изменения численности молоди и взрослых бурых водорослей Ascophyllum nodosum . Mar. Ecol. Прог. Сер. 158 : 111–119. Искать в Google Scholar
Андерссон, С., Л. Каутский и А. Калвас. 1994. Выпуск циркадных и лунных гамет в Fucus vesiculosus в приливной части Балтийского моря. Mar. Ecol. Прог. Сер. 110 : 195–201. Искать в Google Scholar
Ang, P. 1991. Естественная динамика популяции Fucus distichus (Phaeophyceae, Fucales) — воспроизводство и пополнение. Mar. Ecol. Прог. Сер. 78 : 71–85. Искать в Google Scholar
Bäck, S., J.C. Collins and G. Russell. 1991. Аспекты репродуктивной биологии Fucus vesiculosus с побережья юго-запада Финляндии. Офелия 34 : 129–141. Искать в Google Scholar
Bäck, S., Дж. Коллинз и Дж. Рассел. 1993. Сравнительная репродуктивная биология Финского залива и Ирландского моря Fucus vesiculosus L. Sarsia 78 : 265–272. Искать в Google Scholar
Бергер Р., Мальм Т. и Каутский Л. 2001. Две репродуктивные стратегии в Балтике Fucus vesiculosus (Phaeophyceae). Eur. J. Phycol. 36 : 265–273. Искать в Google Scholar
Бергер, Р., Л. Бергстрём, Э. Гран и Л. Каутский. 2004. Как эвтрофикация влияет на разные стадии жизни Fucus vesiculosus в Балтийском море? — концептуальная модель. Hydrobiologia 514 : 243–248. Искать в Google Scholar
Bindoff, N.L., J. Willebrand, V. Artale, A. Cazenave, J. Gregory, S. Gulev, K. Hanawa, C. LeQuéré, S. Levitus, Y. Nojiri, C.K. Шум, Л. Талли и А. Унникришнан. 2007. Наблюдения: изменение климата океана и уровень моря. In : (С. Соломон, Д. Цинь, М. Мэннинг, З. Чен, М. Маркиз, К. Б. Аверит, М. Тиньор и Х. Л. Миллер, ред.) Изменение климата 2007: основы физических наук .Рабочая группа I Четвертого оценочного доклада Межправительственной группы экспертов по изменению климата, Cambridge University Press, Кембридж. С. 129–234. Искать в Google Scholar
Bird, N.L. и Дж. Маклахлан. 1976. Контроль образования цветоложов у Fucus distichus L. subsp. distichus (Phaeophyceae, Fucales). Phycologia 15 : 79–84. Искать в Google Scholar
Brawley, S.H. и Л. Джонсон. 1992. Гаметогенез, гаметы и зиготы: экологическая перспектива полового размножения у водорослей. Br. Phycol. J. 27 : 233–252. Искать в Google Scholar
Brenchley, J.L., J.A. Рэйвен и А. Джонстон. 1996. Сравнение репродуктивного распределения и репродуктивного усилия между семелопущими и итеропородными фукоидами (Fucales, Phaeophyta). Hydrobiologia 326/327 : 185–190. Искать в Google Scholar
Caldeira, K. and M.E. Wickett. 2005. Модели океана предсказывают химические изменения от выбросов углекислого газа в атмосферу и океан. J. Geophys.Res. 110 : 1–12. Искать в Google Scholar
Карлсон Л. 1991. Сезонные колебания в росте, воспроизводстве и содержании азота в Fucus vesiculosu s L. в Эресунде, Южная Швеция. Bot. 34 марта, : 447–453. Искать в Google Scholar
Cousens, R. 1986. Количественное воспроизводство и воспроизводственные усилия насаждениями бурой водоросли Ascophyllum nodosum (L.) Le Jotis на юго-востоке Канады. Estuar. Берег. Shelf Sci. 22 : 495–507.Искать в Google Scholar
Dring, M.J. 1974. Репродукция. In : (W.D.P. Stewart, ed.) Физиология и биохимия водорослей . Ботанические монографии, Univ. Calif. Press, Беркли, Калифорния. С. 814–837. Искать в Google Scholar
Элкен, Дж., А. Леманн и К. Мирберг. 2015. Последние изменения — морская циркуляция и стратификация. In : (Авторская группа BACC II, ред.) Вторая оценка изменения климата для бассейна Балтийского моря . Издательство Springer International Publishing, Гейдельберг, Нью-Йорк, Дордрехт, Лондон.С. 131–144. Искать в Google Scholar
Feely, R.A., C.L. Sabine, K. Lee, W. Berelson, J. Kleypas, V.J. Фабри и Ф.Дж. Миллеро. 2004. Воздействие антропогенного CO 2 на систему CaCO 3 в океанах. Наука 305 : 362–366. Искать в Google Scholar
Gordillo, F.J., F.X. Ниелл и Ф. Фигероа. 2001. Нефотосинтетическое усиление роста за счет высокого уровня CO 2 в нитрофильных морских водорослях Ulva rigida C. Agardh (Chlorophyta). Planta 213 : 64–70. Искать в Google Scholar
Graiff, A., I. Bartsch, W. Ruth, M. Wahl и U. Karsten. 2015a. Сезон оказывает различное воздействие закисления океана и потепления на рост и углеродный метаболизм морских водорослей Fucus vesiculosus в западной части Балтийского моря. Фронт. Mar. Sci. 2 : 1–18. Искать в Google Scholar
Graiff, A., D. Liesner, U. Karsten и I. Bartsch. 2015b. Температурная устойчивость западной части Балтийского моря Fucus vesiculosus — рост, фотосинтез и выживание. J. Exp. Mar. Biol. Ecol. 471 : 8–16. Искать в Google Scholar
Gräwe, U., R. Friedland и H. Burchard. 2013. Будущее западной части Балтийского моря: два возможных сценария. Ocean Dyn. 63 : 901–921. Ищите в Google Scholar
Hales, J.M. and R.L. Fletcher. 1990. Исследования недавно интродуцированной бурой водоросли Sargassum muticum (Yendo) Fensholt. V. Инициирование и рост сосудов, а также высвобождение гамет в лабораторной культуре. Bot. Мар.32 : 167–176. Искать в Google Scholar
HELCOM. 2013. Изменение климата в районе Балтийского моря. Тематическая оценка ХЕЛКОМ в 2013 году. In : Baltic Sea Environment Proceedings No. 137. Helsinki. Искать в Google Scholar
Калвас А. и Каутские Л. 1993. Географические изменения морфологии Fucus vesiculosus в Балтийском и Северном морях. Eur. J. Phycol. 28 : 85–91. Искать в Google Scholar
Каутский Х., Каутский Л., Каутский Н., У.Каутский и К. Линдблад. 1992. Исследования сообщества Fucus vesiculosus в Балтийском море. In : (I. Wallentinus and P. Snoeijs, ред.) Психологические исследования северных прибрежных вод . Acta phytogeogr. Suecica 78: 33–48. Искать в Google Scholar
Kiirikki, M. and A. Lehvo. 1997. Жизненные стратегии нитчатых водорослей собственно Северной Балтики. Сарсия 82 : 259–267. Искать в Google Scholar
Knight, M. and M.Парк. 1950. Биологическое исследование Fucus vesiculosus L. и F. serratus L. J. Mar. Biol. Доц. Соединенное Королевство 29 : 439–501. Искать в Google Scholar
Koch, M., G. Bowes, C. Ross and X.-H. Чжан. 2013. Влияние изменения климата и закисления океана на морские травы и морские макроводоросли. Glob. Чанг. Биол. 19 : 103–132. Искать в Google Scholar
Korpinen, S., T. Honkanen, O. Vesakoski, A. Hemmi, R. Koivikko, J. Loponen and V.Йормалайнен. 2007. Сообщества макроводорослей сталкиваются с проблемой изменения биотических взаимодействий: обзор с акцентом на Балтийское море. Ambio 36 : 203–211. Искать в Google Scholar
Kraufvelin, P., A.T. Руусканен, Н. Наппу и М. Киирикки. 2007. Зимняя колонизация и сукцессия нитчатых макроводорослей на искусственных субстратах и возможные связи с поселением Fucus vesiculosus в начале лета. Estuar. Coast Shelf Sci. 72 : 665–674.Искать в Google Scholar
Kraufvelin, P., A.T. Руусканен, С. Бэк и Г. Рассел. 2012. Повышенная температура морской воды и свет во время ранней весны ускоряют рост вместилищ Fucus vesiculosus в северной части Балтийского моря. Mar. Biol. 159 : 1795–1807. Искать в Google Scholar
Ladah, L.B., F. Feddersen, G.A. Пирсон и Э. Серрао. 2008. Модели высвобождения яиц и расселения двудомных и гермафродитных фукоидных водорослей во время приливного цикла. Mar. Biol. 155 : 583–591. Искать в Google Scholar
Lehvo, A., S. Bäck и M. Kiirikki. 2001. Выращивание Fucus vesiculosus L. (Phaeophyta) в Северной Балтике: хранение энергии и азота в сезонных условиях. Bot. Мар. 44 : 345–350. Искать в Google Scholar
Люнинг, К. и И. Том Дик. 1989. Экологические триггеры сезонности водорослей. Bot. 32 марта, : 389–397. Искать в Google Scholar
Maczassek, K.2014. Экологические факторы плодородия, оплодотворения и прорастания Fucus vesiculosus на побережье Германии. Кандидат наук. диссертация, Христианско-Альбрехтский университет Киля, Киль. Искать в Google Scholar
Maggs, C.A. и M.D. Guiry. 1987. Экологический контроль фенологии макроводорослей. In : (R.M.M. Crawford, ed.) Растительная жизнь в водных и земноводных средах обитания. Специальная публикация № 5 Британского экологического общества, Вили-Блэквелл, Оксфорд. С. 359–373.Ищите в Google Scholar
Mathieson, A.C. and Z. Guo. 1992. Закономерности распределения репродуктивной биомассы фукоидов. Br. Phycol. J. 27 : 271–292. Искать в Google Scholar
Mathieson, A.C., J.W. Шипман, Дж. Р. О’Ши и Р.С. Хашевлат. 1976. Сезонный рост и размножение эстуарных фукоидных водорослей в Новой Англии. J. Exp. Mar. Biol. Ecol. 25 : 273–284. Искать в Google Scholar
Middelboe, A.L., K. Sand-Jensen and T. Binzer. 2006. Высоко предсказуемая фотосинтетическая продукция в естественных сообществах макроводорослей за счет входящего и поглощенного света. Oecologia 150 : 464–476. Искать в Google Scholar
Nygård, C.A. и М.Дж. Дринг. 2008. Влияние солености, температуры, растворенного неорганического углерода и концентрации питательных веществ на фотосинтез и рост Fucus vesiculosus из Балтийского и Ирландского морей. Eur. J. Phycol. 43 : 253–262. Искать в Google Scholar
Olischläger, M. and C. Wiencke. 2013. Закисление океана смягчает воздействие низких температур на рост и фотосинтез красной водоросли Neosiphonia harveyi (Rhodophyta). J. Exp. Бот. 64 : 5587–5597. Искать в Google Scholar
Olischläger, M., I. Bartsch, L. Gutow и C. Wiencke. 2012. Влияние закисления океана на различные стадии жизненного цикла ламинарии Laminaria hyperborea (Phaeophyceae). Bot. 55 марта, : 511–525. Искать в Google Scholar
Orr, J.C., V.J. Фабри, О. Омон, Л. Бопп, С.С. Дони, Р.А. Feely, A. Gnanadesikan, N. Gruber, A. Ishida, F. Joos, R.M. Ки, К. Линдси, Э. Майер-Реймер, Р. Матеар, П.Monfray, A. Mouchet, R.G. Наджар, Г.-К. Платтнер, К. Роджерс, К. Sabine, J.L. Sarmiento, R. Schlitzer, R.D. Slater, I.J. Тоттерделл, М.-Ф. Weirig, Y. Yamanaka и A. Yool. 2005. Антропогенное закисление океана в XXI веке и его влияние на кальцифицирующие организмы. Природа 437 : 681–686. Искать в Google Scholar
Pearson, G.A. и С. Броули. 1996. Репродуктивная экология Fucus distichus (Phaeophyceae): литоральная водоросль с успешным внешним оплодотворением. Mar. Ecol. Прог. Сер. 143 : 211–223. Искать в Google Scholar
Pearson, G.A. и Э.А. Серрао. 2006. Возвращаясь к вопросу о синхронном высвобождении гамет фукоидными водорослями в приливной зоне: успех оплодотворения и не только? Integr. Комп. Биол. 46 : 587–597. Искать в Google Scholar
Pearson, G.A., E.A. Серрао и С. Броули. 1998. Контроль высвобождения гамет у фукоидных водорослей: определение гидродинамических условий посредством поглощения углерода. Экология 79 : 1725–1739.Искать в Google Scholar
Pierrot, D., E. Lewis and D.W.R. Уоллес. 2006. Разработана программа MS Excel для расчетов системы CO 2 . В : ORNL / CDIAC-105a. Национальная лаборатория Ок-Ридж, Министерство энергетики США, Теннесси. Искать в Google Scholar
Provasoli, L. 1968. СМИ и перспективы выращивания морских водорослей. В : (А. Ватанабе и А. Хаттори, ред.) Культуры и коллекции водорослей . Труды американо-японской конференции, Хаконэ, 1966 г.Японское общество физиологии растений, Токио. С. 63–75. Ищите в Google Scholar
R Development Core Team. 2016. R: язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. URL: http://www.R-project.org/. Искать в Google Scholar
Råberg, S., R. Berger-Jönsson, A. Björn, E. Granéli и L. Kautsky. 2005. Воздействие Pilayella littoralis на пополнение Fucus vesiculosus : последствия для состава сообщества. Mar. Ecol. Прог. Сер. 289 : 131–139. Искать в Google Scholar
Robertson, B.L. 1987. Репродуктивная экология и структура полога Fucus spiralis L. Bot. 30 марта, : 475–482. Искать в Google Scholar
Rönnbäck, P., N. Kautsky, L. Pihl, M. Troell, T. Söderqvist и H. Wennhage. 2007. Экосистемные товары и услуги из шведских прибрежных местообитаний: идентификация, оценка и последствия экосистемных сдвигов. Ambio 36 : 534–544.Искать в Google Scholar
Ruuskanen, A. and S. Bäck. 1999. Влияет ли экологический стресс на плодовитость и регенерацию вайя Fucus vesiculosus ? Finnish Zool. Бот. Publ. Доска 36 : 285–290. Искать в Google Scholar
Sabine, C.L., R.A. Фили, Н. Грубер, Р. Key, K. Lee, J.L. Bullister, R. Wanninkhof, C.S. Wong, D.W.R. Уоллес, Б. Тилбрук, Ф.Дж. Миллеро, Т.-Х. Пэн, А. Козырь, Т. Оно и А.Ф. Риос. 2004. Океанический сток антропогенного CO 2 . Наука 305 : 367–371. Искать в Google Scholar
Saderne, V. 2012. Экологическое воздействие CO 2 на бурые водоросли Fucus serratus и его эпибионты: от среды обитания до масштаба организма. Кандидат наук. диссертация, Христианско-Альбрехтский университет Киля, Киль. Искать в Google Scholar
Santelices, B. 1990. Модели размножения, распространения и пополнения у морских водорослей. В : (М. Барнс, ред.) Oceanogr. Mar. Biol. Анну. Rev. 28: 177–276.Искать в Google Scholar
Sarker, M.Y., I. Bartsch, M. Olischläger, L. Gutow и C. Wiencke. 2013. Комбинированное воздействие CO 2 , температуры, освещенности и времени на физиологические характеристики Chondrus crispus (Rhodophyta). Bot. 56 марта, : 63–74. Искать в Google Scholar
Schiel, D.R. 1985. Краткосрочное демографическое исследование Cystoseira osmundacea (Fucales: Cystoseiraceae) в центральной Калифорнии. J. Phycol. 21 : 99–106.Искать в Google Scholar
Шнайдер Б., К. Эйлола, К. Луккари, Б. Мюллер-Карулис и Т. Нойман (2015) Воздействие на окружающую среду — Морская биогеохимия. In : (Авторская группа BACC II, ред.) Вторая оценка изменения климата для бассейна Балтийского моря . Издательство Springer International Publishing, Гейдельберг, Нью-Йорк, Дордрехт, Лондон. С. 337–361. Искать в Google Scholar
Serrão, E.A., L. Kautsky and S.H. Броули. 1996a. Успех распространения морских водорослей Fucus vesiculosus L.в солоноватом Балтийском море коррелирует с осмотическими возможностями балтийских гамет. Oecologia 107 : 1–12. Искать в Google Scholar
Serrão, E.A., G. Pearson, L. Kautsky and S.H. Броули. 1996b. Успешное внешнее оплодотворение в турбулентной среде. Proc. Natl. Акад. Sci. США 93 : 5286–5290. Искать в Google Scholar
Serrão, E.A., S.H. Броули, Дж. Хедман и Л. Каутский. 1999. Успешное воспроизводство Fucus vesiculosus (Phaeophyceae) в Балтийском море. J. Phycol. 35 : 254–269. Искать в Google Scholar
Terry, L.A. and B.L. Мох. 1980. Влияние фотопериода на инициацию сосудов у Ascophyllum nodosum (L.) Le Jol. Br. Phycol. J. 15 : 291–301. Искать в Google Scholar
Торн, К., Д. Краузе-Йенсен и Г. Мартин. 2006. Распределение мочевого пузыря ( Fucus vesiculosus, ) в Балтийском море по глубине в настоящее время и в прошлом. Aquat. Бот. 84 : 53–62. Искать в Google Scholar
Underwood, A.J. 1997. Эксперименты в области экологии: их логический план и интерпретация с использованием дисперсионного анализа . Издательство Кембриджского университета, Кембридж. С. 140–197. Искать в Google Scholar
Vernet, P. and J. Harper. 1980. Цена секса в водорослях. Biol. J. Linn. Soc. 13 : 129–138. Искать в Google Scholar
Wahl, M., V. Jormalainen, B.K. Эрикссон, Дж. Койер, М. Молис, Х. Шуберт, М. Детье, Р. Карез, И. Круз, М. Ленц, Г. Пирсон, С. Роде, С.А. Викстрём и Дж.Л. Ольсен. 2011. Экология стресса у фукусов: абиотические, биотические и генетические взаимодействия. В : (М. Лессер, ред.) Успехи в морской биологии . Elsevier Ltd, Оксфорд. С. 37–105. Искать в Google Scholar
Wahl, M., M. Molis, A.J. Hobday, S. Dudgeon, R. Neumann, P. Steinberg, A.H. Campbell, E. Marzinelli и S. Connell. 2015a. Реакция бурых макроводорослей на изменение окружающей среды от локального к глобальному масштабу: прямые и экологически опосредованные эффекты. Перспектива.Phycol. 2 : 11–30. Искать в Google Scholar
Wahl, M., B. Buchholz, V. Winde, D. Golomb, T. Guy-Haim, J. Müller, G. Rilov, M. Scotti, M.E. Böttcher. 2015b. Концепция мезокосма для моделирования почти естественного неглубокого подводного климата: Kiel Outdoor Benthocosms (KOB). Лимнол. Oceanogr. Методы 13 : 651–663. Искать в Google Scholar
Warton, D.I. и F.K.C. Хуэй. 2011. Арксинус глуп: анализ пропорций в экологии. Экология 92 : 3–10.Искать в Google Scholar
Wu, H., D. Zou and K. Gao. 2008. Воздействие повышенной концентрации CO в атмосфере 2 на фотосинтез и рост микро- и макроводорослей. Sci. China Ser. C Life Sci. 51 : 1144–1150. Искать в Google Scholar
Молекул | Бесплатный полнотекстовый | Ранний эмбриогенез бурой водоросли Fucus vesiculosus L. характеризуется значительными изменениями углеродного и энергетического метаболизма
3.1. Общие соображения
Это исследование представляет первую характеристику динамического метаболического профиля во время фукоидного эмбриогенеза.Список метаболитов, идентифицированных в эмбрионах фукуса (Таблица S1), в целом хорошо согласуется с результатами предыдущих биохимических отчетов о бурых водорослях [26,29,33,35,36,37,38], но также содержит соединения, описанные для впервые в фукоидах. Самым распространенным растворимым углеводом является маннит, тогда как другие свободные сахара и полиолы (глюкоза, фруктоза, сахароза, мио-инозит) присутствуют в значительно меньших количествах. Скорее всего, это результат их быстрого превращения в запасные соединения, такие как маннит и ламинаран, через гексозо-фосфаты (рис. 4).Преобладающими органическими кислотами являются цитрат и малат (рис. 5, таблица S1). Детальное исследование, проведенное на Ectocarpus [26], также показало высокий уровень цитрата, но неопределяемое количество малата. Мы предполагаем, что то же самое может быть и для взрослых талломов F. vesiculosus, где заключительные стадии раннего эмбриогенеза сопровождаются накоплением цитрата и уменьшением количества малата (Рисунок 3 и Рисунок 5). В клетках растений малат участвует в анаплеротических реакциях в митохондриях, восполняя потерю цитрата и α-кетоглутарата из цикла TCA для биосинтеза аминокислот и жирных кислот.Малат окисляется до пирувата, образуя НАДН с помощью НАД-яблочного фермента в митохондриальном матриксе, что позволяет цитрату образовываться без полного цикла TCA. Вероятно, накопленный пул малата в яйцах фукоидов способствует активным процессам биосинтеза в первые дни эмбриогенеза. Среди свободных протеиногенных аминокислот, обнаруженных в яйцах, зиготах и эмбрионах F. vesiculosus, наиболее распространены аланин, глутамат, аспартат и серин (Таблица S1). Как правило, аминокислотные профили соответствуют литературным данным для различных бурых водорослей и напоминают профили, наблюдаемые у высших растений [26,39].Помимо обычных первичных метаболитов растительных клеток, профили F. vesiculosus содержат большое количество сахаров и полиолов с относительно высокой молекулярной массой, которые обычно трудно однозначно идентифицировать (Таблица S1). Скорее всего, большинство из них представляет собой различные продукты гидролиза соединения длительного хранения углерода ламинарана, представляющего собой полидисперсный полимер на основе β-1,3-глюкозы, содержащий до 3,7% маннита [40,41].
3.2. Развитие зиготы
Многие ключевые физиологические процессы происходят во время одноклеточной стадии фукоидного эмбриогенеза (первые ~ 20 ч AF).Специфика этой стадии развития отражена в результатах PCA наших данных профилирования метаболитов, где первый главный компонент отчетливо объединяет все оценки, связанные с яйцами / зиготами (до 12 ч AF), отделяющими их от эмбрионов (Рисунок 2). Наиболее значимыми физиологическими событиями на этой стадии являются: оплодотворение яйцеклетки, начальный синтез клеточной стенки в зиготе, прикрепление к субстрату, поляризация клеток в соответствии с сигналами окружающей среды и образование ризоидного выступа (прорастание зиготы) [14,15].Наши данные показывают, что биохимические процессы, лежащие в основе этих событий, приводят к изменениям в содержании соединений, связанных с дыханием и метаболизмом липидов и аминокислот (Рисунок 2 и Рисунок 3, Таблица 1). Неоплодотворенные яйца фукоидов могут выжить в морской воде в течение нескольких дней и не менее 50% из них могут быть успешно оплодотворены в течение 48 часов после выхода из оогониев (данные не показаны). Они уже могут фотосинтезировать, хотя интенсивность фотосинтеза намного меньше, чем у многодневных эмбрионов [7,20,42].Эти ограниченные фотосинтетические способности и потенциальная способность фукоидных эмбрионов развиваться до некоторой степени в темноте подразумевают, что запасные соединения, сконцентрированные в яйцеклетках, должны быть основным источником углерода и энергии во время раннего эмбриогенеза [43,44]. Двумя наиболее важными соединениями, запасающими углерод в бурых водорослях, являются маннит сахарного спирта, который также является основным стабильным продуктом фотосинтеза, и полимерный ламинаран β-1,3-глюкозы [25,40]. Хотя подробный механизм ремобилизации ламинарана в водорослях все еще обсуждается, данные профилирования метаболитов подтверждают, что ключевым событием первого часа AF является отвлечение углеродных потоков от ламинарана, маннита и других полиолов и сахаров (фруктозы, арабинозы и т. дисахариды) в более метаболически активные вещества — сахар-фосфаты и глицерин (Рисунок 3 и Рисунок 4), что является широко принятым понятием для ремобилизации ламинарана.Следовательно, содержание ламинарана снижается в 5 раз в течение первого часа развития зиготы (Рисунок 4) — этот результат согласуется с данными Quatrano и Stevens (1976), которые сообщили о снижении ламинарана через 7 часов AF [43]. Относительное изменение концентрации маннита не такое большое (хотя и статистически значимое), скорее всего, потому, что это вещество также используется в качестве совместимого растворенного вещества и не должно резко меняться для поддержания постоянных осмотических условий [45]. Однако Allen et al. (1972) показали значительное увеличение внутреннего осмотического давления в только что оплодотворенных фукоидных зиготах, что могло быть результатом притока K + и Cl — [23].В течение первых трех часов после оплодотворения образуется клеточная стенка, и, таким образом, яйца обязательно находятся в осмотическом равновесии с морской водой или близком к нему. После этого внутреннее осмотическое давление быстро увеличивается и к началу прорастания достигает равновесия. Наши данные также предполагают, что накопление глицерина может способствовать поддержанию раннего осмотического равновесия с морской водой до начала притока K + и Cl —, особенно во время начальной фазы формирования клеточной стенки.Общее снижение содержания сахара и полиола совпадает с накоплением веществ, связанных с дыханием (рис. 3), и известно, что интенсивность дыхания значительно увеличивается в первые часы фукоидного эмбриогенеза [46,47]. Однако данные профилирования метаболитов также показывают более выраженное изменение содержания сахар-фосфатов (фруктозо-6-фосфат, глюкозо-6-фосфат, мио-инозитол-1-фосфат), чем в содержании промежуточных продуктов цикла TCA, которые являются в основном довольно стабильный или слегка снижающийся в течение первого часа AF, и только фумаровая кислота демонстрирует значительное увеличение (Рисунок 3 и Рисунок 5).Это может быть результатом учащенного дыхания, когда метаболиты цикла TCA не накапливаются из-за их высокой потребности. Также это может означать, что значительная часть сахаров-фосфатов расходуется не на дыхание, а на биосинтетические реакции. Ранее было показано, что эти соединения находятся на перекрестке различных метаболических путей, являясь ключевыми элементами центрального метаболизма углерода у бурых водорослей [25]. В тканях талломов взрослых F. vesiculosus сахар-фосфаты, глицерин и глицериновая кислота являются основными метаболитами, способствующими быстрому распределению углерода [48].Путь, ведущий к образованию альгината и сульфатированных фуканов, начинается с фруктозо-6-фосфата [49,50], в то время как глюкозо-6-фосфат может потребляться для биосинтеза целлюлозы [51], а мио-инозитол-1-фосфат является исходным предшественником. различных полисахаридов клеточной стенки [52]. Все вместе эти биосинтетические процессы обеспечивают материалы для формирования клеточной стенки и отложения стенок на поверхности фукоидных зигот, начиная с нескольких минут после оплодотворения [8]. Было показано, что альгинаты являются первыми составляющими стенки, появляющимися на поверхности клетки, и целлюлоза может быть обнаружена в течение ~ 20 мин AF, а сульфатированные фуканы — в течение 1 часа AF [43,53].Другой процесс, который, по-видимому, начинается параллельно с отложением клеточной стенки, — это выделение адгезивного материала, состав которого аналогичен матрице стенки [11]. Прикрепление к субстрату — одна из самых актуальных задач развития зигот, необходимое условие восприятия поляризующих раздражителей окружающей среды [9,11]. Оба процесса, интенсивное осаждение клеточной стенки и образование адгезива, продолжаются до ~ 6 часов AF, когда состав клеточной стенки становится стабильным и происходит необратимое затвердевание адгезивного вещества [11,43,53].Помимо предшественников клеточной стенки и адгезивных компонентов, эта усиленная везикулярная секреция требует пула мембранных компонентов. Данные профилирования метаболитов показывают постоянное увеличение количества соединений, связанных с метаболизмом липидов, в течение первых 6 часов развития зигот (Рисунок 3 и Рисунок 5). Наблюдаемое несоответствие в динамике компонентов цикла TCA — обратное поведение лимонной кислоты и дикарбоновых кислот (рис. 3) — также может быть результатом интенсивного липидного обмена в первые часы эмбриогенеза Fucus.Лимонная кислота является одним из центральных метаболитов в биохимии растений, опосредуя перекрестное взаимодействие различных путей, таких как цикл TCA, цикл глиоксилата и биосинтез липидов. Таким образом, снижение содержания лимонной кислоты в течение 1 часа AF, сохраняющееся в течение следующих 2 часов в контексте относительно повышенных уровней концентрации других метаболитов TCA, может быть связано с ее потреблением для биосинтеза жирных кислот и сквалена, а затем с образованием мембран (рис. Как только фукоидные зиготы прикрепляются к субстрату, они становятся чувствительными к серии сигналов окружающей среды, в соответствии с которыми они формируют свою начальную ось полярности и начинают процесс дифференцировки [12].Поскольку дифференцировка зиготы (в первую очередь — инициация роста кончика, которая происходит примерно через 12 часов AF) также требует активного везикулярного транспорта на основе компонентов мембраны [8,54], мы предполагаем, что после 6 часов развития, когда формирование клеточной стенки в основном происходит готово, это будет основной приемник жирных кислот и сквалена. Таким образом, резкое увеличение количества соединений, связанных с метаболизмом липидов, в период около 6 часов AF (Рисунок 3 и Рисунок 5), по-видимому, поддерживает все три процесса — отложение клеточной стенки, образование адгезивов и дифференцировку зигот.Quatrano (1978) показал, что белки, необходимые для первого деления зиготы и образования ризоида, синтезируются прибл. между 8 и 13 часами AF [12]. Синтез мРНК de novo начинается в фукоидных зиготах в течение часа AF, что является предпосылкой для последующих процессов поляризации и роста [12,14]. Это совпадает с увеличением содержания аминокислот в первые часы развития зиготы (рис. 3). Высокая потребность метаболита TCA α-кетоглутарата для биосинтеза аминокислот также может способствовать низкому уровню цитрата на этой стадии.Мы с трудом могли обнаружить α-кетоглутарат в профилях метаболитов эмбриона — скорее всего, из-за его быстрого потребления. Временное падение концентрации большинства аминокислот в период между 3 и 6 часами AF может быть результатом их массового потребления для биосинтеза белка, поддерживающего основные события этой эмбриогенной стадии. Только две аминокислоты, а именно глицин и серин, значительно увеличиваются в этот период. Общая особенность этих двух аминокислот состоит в том, что они обе участвуют в фотодыхании.Предположительно появление полноценной клеточной стенки заставляет зиготу перестраивать свой метаболизм, поскольку стенка препятствует газообмену и диффузии гликолата, что приводит к активации реакций фотодыхания. Таким образом, период около 6 часов AF может быть временем адаптации клетки к изменившимся скоростям диффузии. Через 12 часов AF происходит активация большинства аминокислот, по-видимому, для поддержки новой волны интенсификации синтеза белка.
3.3. Эмбриогенез после первого деления зиготы
Согласно анатомо-физиологическим данным, первые дни фукоидного эмбриогенеза после деления зиготы включают такие основные процессы, как повышение эффективности фотосинтеза, интенсивное удлинение ризоидов и начало органогенеза [17,19,20].Данные профилирования метаболитов показывают, что наиболее заметное различие между зиготами и эмбрионами заключается в содержании продуктов фотосинтеза — сахаров и полиолов — начиная с 1 дня AF (Рисунок 3 и Рисунок 4). Значительно увеличивается концентрация маннита, а также различных моно-, ди- и трисахаридов. Известно, что фотосинтетическая активность фукоидных зародышей быстро возрастает в первые дни эмбриогенеза [7,20,42]. Максимальная скорость транспорта электронов у зародышей F. vesiculosus увеличивается более чем в два раза в течение первых трех дней развития [20].Мы можем сделать вывод, что накопление сахара и полиола, начинающееся вскоре после первого деления зиготы, означает, что фотосинтез стал достаточно эффективным, чтобы восполнить энергетические и материальные затраты и инициировать накопление ресурсов для последующего развития. Содержание соединений, связанных с дыханием, за исключением цитрата, обычно увеличивается. вниз, предположительно отражая повышение эффективности фотосинтеза (Рисунок 3 и Рисунок 5). Всплеск сахарофосфатов на 3-й день хорошо совпадает с периодом быстрого удлинения ризоида за счет роста апикальных кончиков клеток.Рост кончика носа — это процесс, сильно зависящий от энергии и субстрата, основанный на постоянном производстве и перераспределении множества компонентов мембраны и клеточной стенки [55]. Поскольку нет синхронного увеличения других соединений, связанных с дыханием, этот сахарно-фосфатный всплеск, скорее всего, связан с интенсификацией биосинтетических процессов, поддерживающих удлинение ризоидов. Согласно нагрузкам PCA, фенольные соединения представляют собой еще одну группу метаболитов, вносящих существенный вклад в разницу между стадиями эмбриогенеза Fucus (табл. 1).Наблюдается общее падение фенольных соединений в течение 6–12 часов с последующим накоплением в 3–6-дневных эмбрионах с выбросом флороглюцина у однодневных эмбрионов (рис. 3). Поскольку вторичный метаболизм бурых водорослей все еще плохо изучен, эта часть результатов наиболее интересна, но ее трудно интерпретировать. Флороглюцин является мономером особой группы фенольных соединений бурых водорослей — флортанинов [56]. Предполагаемый спектр физиологических функций этих полифенолов включает антиоксидантную активность и химическую защиту от биообрастания и выпаса скота [22,57].Кроме того, они являются неотъемлемыми компонентами стенок фукоидных ячеек и адгезивного материала [53,58]. Хотя состав клеточной стенки стабилизируется до 6 часов AF, ее толщина все еще увеличивается до 12 часов AF [43], что означает постоянное потребление компонентов стенки. Полифенолы откладываются преимущественно в стенке ризоидного выступа прорастающей зиготы, таким образом, являясь одним из последних компонентов, включенных в стенку зиготической клетки [59]. Таким образом, постепенное уменьшение свободного флороглюцина в течение 1–12 ч AF (рис. 3) могло быть результатом его быстрой полимеризации и включения в клеточную стенку.Временное накопление фенольных соединений через 12 часов AF, вызывающее всплеск на 1 день AF, может быть признаком завершения формирования клеточной стенки. Синхронные изменения в содержании флороглюцина и производного флороглюциновой кислоты, последнее с его наименьшим значением предшествует наивысшей концентрации флороглюцина, с последующей активацией двух других обнаруженных фенольных соединений в течение этого периода эмбриогенеза, подразумевает, что эти соединения должны быть метаболически связаны. Однако до настоящего времени пути биосинтеза флоротаннина были мало изучены на биохимическом и молекулярном уровнях [60], и возможная роль флороглюциновой кислоты, будь то побочный продукт, предшественник или продукт флороглюцина, требует дальнейшего изучения.Флороглюциновая кислота и ее метиловый эфир были ранее выделены из нескольких высших растений [61], но, насколько нам известно, это первый намек на потенциальное присутствие этого соединения в бурых водорослях. Как правило, через 3 дня развития метаболические профили эмбрионы все больше напоминают зародыши взрослых бурых водорослей. Содержание аминокислот, жирных кислот и сквалена значительно снижается (рисунок 3 и рисунок 5). Среди органических кислот цикла TCA наблюдается значительное накопление лимонной кислоты, что особенно заметно у 6–9-дневных эмбрионов.Эта органическая кислота имеет тенденцию доминировать в метаболических профилях взрослых бурых водорослей [26], включая фукоиды [38]. Предположительно, его низкий уровень в зиготах и молодых эмбрионах является специфической биохимической особенностью этой стадии развития, когда лимонная кислота массово расходуется на биосинтетические реакции, такие как образование мембран и биосинтез аминокислот.
Volume 65 No 2 page 109
ЭКСТРАКТ FUCUS: КОСМЕТИЧЕСКОЕ ЛЕЧЕНИЕ ДЛЯ ТЕМНЫХ КРУГОВ ПОД ГЛАЗАМИ 109 Результаты были дополнительно подтверждены исследованием зависимости от дозы в кератиноцитах, как показано на рисунке 3.В концентрациях от 0,5% до 2% экстракт фукуса положительно стимулировал экспрессию гена HO-1 в культуре кератиноцитов. При 1% экстракт фукуса стимулировал HO-1 до 2,2 log, что более чем в 4 раза превышает контроль при 2%, стимуляция пошла примерно в 10 раз по сравнению с контролем (p 0,001). Стимулирующая активность экстракта фукуса HO-1 на уровне белка, проанализированная с помощью вестерн-блоттинга, представлена на Фигуре 4 для кератиноцитов и на Фигуре 5 для клеток фибробластов.Экстракт фукуса в концентрациях 1% и 2% индуцировал белок HO-1 в кератиноцитах в значительной степени дозозависимым образом (рис. 4). Количественный денситометрический анализ интенсивности полос белка показал, что при 1% экстракт фукуса индуцировал белок HO-1 примерно в 6 раз по сравнению с контролем, при 2% он индуцировал белок HO-1 примерно в 30 раз по сравнению с контролем. Значения p парного t-критерия Стьюдента для обеих концентраций меньше 0,001 по сравнению с контролем, что является статистически значимым.В клетках фибробластов (рис. 5) белок HO-1 был индуцирован экстрактом фукуса при концентрациях 1% и 2%. Когда интенсивность полос анализируемого белка сравнивалась с контролем для парного t-критерия Стьюдента, значения p для обеих концентраций были меньше 0,01, что является статистически значимым. Индуцированное содержание белка HO-1 в клетках фибробластов достигало максимального уровня при обработке 1% экстракта фукуса. Эти результаты ясно показывают, что экстракт фукуса способен индуцировать HO-1 как в клетках кератиноцитов, так и в клетках фибробластов, что позволяет предположить, что экстракт фукуса может иметь полезные свойства. тип NHDF NHEK HO-1 log2-кратное изменение относительно контроля 1.6 5,8 НО-1-кратное изменение относительно контроля 3 55 Рис. 4. Экстракт фукуса увеличил содержание НО-1 в кератиноцитах по данным вестерн-блоттинга. Среднее значение ± стандартное отклонение по 3 анализам — парный t-критерий Стьюдента — ***: p 0,001.
ЖУРНАЛ КОСМЕТИЧЕСКОЙ НАУКИ 110 уменьшает темные круги под глазами, вызванные утечкой крови и накоплением гема. ВЛИЯНИЕ ЭКСТРАКТА FUCUS НА АНТИОКИСЛЕНИЕ С ПОМОЩЬЮ DPPH Антиоксиданты помогают предотвратить и восстановить повреждение тканей вашего тела, замедляя или предотвращая действие свободных радикалов, а также помогают защитить вашу кожу от разрушительного воздействия солнца (21).DPPH — это обычное сокращение для органического химического соединения 2,2-дифенил-1-пикрилгидразил. Это кристаллический порошок темного цвета, состоящий из стабильных молекул свободных радикалов. Он используется в обычном анализе для определения антиоксидантной способности активных косметических ингредиентов. Рисунок 5. Экстракт фукуса увеличил содержание HO-1 в фибробласте по данным вестерн-блоттинга. Среднее значение ± стандартное отклонение по 3 анализам — парный t-критерий Стьюдента — **: p 0,01 — ***: p 0.001. Фигура 6. Экстракт фукуса: антиоксидантный эффект Среднее + стандартное отклонение трех анализов — парный t-критерий Стьюдента — ***: p 0,001.
прочтений: протисты | Лабораторное руководство Biology II
Цель этого упражнения — узнать о простейших. Мы будем изучать основные группы, а для большинства групп мы будем изучать репрезентативные роды.
Процедура
За каждый образец:
- Внимательно прочтите информацию.
- Создавайте заметки. Ваши заметки будут наиболее полезными, если они будут включать рисунок, описание и важную информацию, такую как жизненный цикл, коммерческая ценность, экологическая значимость и необычные характеристики.
Эвгленозойские
Кинетопластиды
Кинетопластиды жгутиковые, одноклеточные. У них есть темная область митохондрий, которая называется кинетопласт.
Некоторые кинетопластиды находятся в симбиотических (близких) отношениях с другими организмами.Трипаносомы — это кинетопластиды, вызывающие африканскую сонную болезнь. Они передаются своим хозяевам через укус мухи цеце. Трипаносома вызывает африканскую сонную болезнь.
Эвглена
Эвглены одноклеточные. Многие эвглениды питаются фагоцитозом. Многие виды эвгленид фотосинтезируют, но могут стать гетеротрофными при отсутствии солнечного света (миксотрофы).
Euglena использует жгутики для передвижения. Внешнее покрытие, называемое пленкой , является гибким и помогает двигаться.У некоторых есть глазное пятно с фоторецептором, способным обнаруживать присутствие света. Размножение бесполое.
Диатомовые водоросли
Диатомовые водоросли — самые многочисленные одноклеточные водоросли в океанах и, как таковые, являются важным источником пищи и кислорода. Они также важны в пресноводных средах. Они улавливают от 20 до 25% солнечной энергии, улавливаемой живыми организмами. Стенки клеток диатомовых водорослей содержат кремнезем (компонент стекла) и состоят из двух половинок, как дот. Их остатки образуют кизельгур.Он используется для фильтрующих средств и абразивов, таких как чистящие порошки. Диатомовые водоросли являются основным компонентом фитопланктона в пресноводных и морских средах.
Бурые водоросли
Бурые водоросли — автотрофы (фотосинтетические). Их характерный коричневый цвет обусловлен каротиноидными пигментами. Они многоклеточные и имеют размер от маленького до очень большого. Некоторые из них имеют длину от 50 до 100 метров. Их часто можно найти на скалистых берегах в умеренном климате. Тело ( слоевища ) содержит фиксаторов для прикрепления, лопастей , а стержнеобразная структура, удерживающая лопасти, называется ножкой .У многих видов есть поплавки, которые работают в режиме плавучести. У некоторых есть газонаполненные поплавки. Слизистый (слизистый) материал в стенках клеток замедляет высыхание у подвергшихся воздействию людей при отливе. Большинство видов имеют жизненный цикл с сменой поколений .
Фукус
Фукус — это обычная «водоросль», встречающаяся на скалистом побережье. Некоторые виды Fucus имеют диплоидных взрослых особей .
Рис. 1. Гаметы образуются в цветоложах.
Macrocystis и Nereocystis
Macrocystis и nereocystis — это глубоководные водоросли.
Саргассам
Саргассам иногда отламывается, образуя плавучие массы. Вокруг этих масс собираются другие морские организмы.
Ламинария
Ламинария — это коричневая водоросль, которая обычно прикрепляется чуть ниже приливной зоны. Имеет жизненный цикл со сменой поколений.
Рисунок 2. Смена поколений
Динофлагелляты
Защитные пластины из целлюлозы покрывают динофлагелляты, а два жгутика позволяют им двигаться.Один из жгутиков расположен в поперечной бороздке, которая заставляет клетку вращаться при движении.
Большинство из них обитает в морской или пресноводной среде, и многие из них являются фотосинтетическими. Они являются важными компонентами фитопланктона и, следовательно, важны в водных пищевых цепях . В эту группу также входят многие гетеротрофные и многие миксотрофные виды.
Некоторые виды ответственны за красные приливы, убивающие рыбу и моллюсков (Gymnodinium, Gonyaulax, Pfiesteria).Некоторые из них живут как симбиантов внутри некоторых беспозвоночных. Например, некоторые кораллы растут быстрее, поскольку в их клетках живут динофлагелляты. Некоторые виды способны к биолюминесценции (излучают свет).
Происходит как половое, так и бесполое размножение. При половом размножении образуются цисты, устойчивые к неблагоприятным условиям окружающей среды. Кисты бездействуют и становятся активными при улучшении условий окружающей среды.
Инфузории
К этой группе принадлежит род Vorticella .
Парамеций
Рис. 3. Paramecium caudatum X 100
Пелликула (внешнее покрытие) парамеции покрыта сотнями ресничек . У них есть множество органелл, включая пищевод (оральную бороздку) и анальную пору. Инфузории имеют крупное макроядро и более мелкое микроядро.
Микронуклеус участвует в половом и бесполом размножении. Другой ядерной деятельностью занимается макронуклеус. Макронуклеус — это полиплоид (приблизительно 860 N в Paramecium aurelia), а микроядро — это диплоид .
Рисунок 4. Парамеций X 200
Во время размножения макронуклеус распадается. Позже микронуклеус разовьется в макронуклеус. В большинстве случаев размножение происходит бесполым путем (митоз). Половое размножение — спряжение .
Микроядро делится мейозом; 3 из 4 образовавшихся ядер распадутся, как и макронуклеус. Оставшееся гаплоидное ядро разделится путем митоза, в результате чего появится особь с двумя гаплоидными ядрами. Каждый из двух конъюгированных особей обменивается одним из ядер.Затем два гаплоидных ядра сливаются, образуя диплоидное ядро.
Красные водоросли
Красные водоросли в основном многоклеточные и встречаются в основном в более теплых тропических океанах. Их красный цвет обусловлен дополнительным фотосинтетическим пигментом , который называется фикоэритрин. Дополнительные пигменты красных водорослей способны поглощать синий и зеленый свет. Это позволяет некоторым видам выживать в глубоких водах, где преобладает синий и зеленый свет.
Некоторые виды нитчатые , но большинство из них имеют сложный образец ветвления.Некоторые формы кораллина откладывают карбонат кальция в своих клеточных стенках, что способствует развитию коралловых рифов.
Зеленые водоросли
Четыре распространенные формы зеленых водорослей — одноклеточные, колониальные , нитчатые и многоклеточные . Зеленые водоросли считаются предками первых растений. Оба вида организмов имеют следующие общие характеристики:
- У них есть клеточная стенка , содержащая целлюлозу .
- У них хлорофиллов, а и б.
- Они хранят свою пищу как крахмал внутри хлоропласта .
Большинство видов пресноводных, но есть много морских видов. Некоторые живут во влажной почве.
Хламидомонада
Хламидомонада — одноклеточный организм с двумя жгутиками . Хотя этот организм представляет собой единственную клетку, жизненный цикл аналогичен таковому у гаплоидных взрослых взрослых .
Рисунок 5.Жизненный цикл хламидомонады
Размножается бесполым путем (митозом) при благоприятных условиях. Половое размножение происходит в неблагоприятных условиях. Зигота образует толстостенную зигоспору, которая устойчива к экстремальным условиям окружающей среды и делится мейозом, когда условия окружающей среды становятся благоприятными.
Большинство видов Chlamydomonas изогамны (обе гаметы одинакового размера; они изогаметы), некоторые — оогамны (гаметы двух размеров; большие гаметы — яйца, меньшие — сперматозоиды).
Volvox
Volvox — колониальные зеленые водоросли. Клетки расположены в студенистой сфере с двумя жгутиками, направленными наружу. Они делятся бесполым путем, образуя дочернюю колонию.
Некоторые клетки специализируются на производстве сперматозоидов и яйцеклеток для полового размножения. Специализация клеток, наблюдаемая в репродуктивных клетках, характерна для многоклеточных организмов. Вольвокс считается колонией, потому что он занимает промежуточное положение между группой отдельных клеток и многоклеточным организмом.
Спирогира
Спирогира представляет собой нитевидную форму . Имеет ленточный спиралевидный хлоропласт. В жизненном цикле есть гаплоидные взрослые особи .
Половое размножение происходит путем спряжения . Конъюгация относится к процессу, при котором гаметы передаются от одного человека к другому посредством связи между ними.
Зигота устойчива и зимует. Весной он делится мейозом с образованием гаплоидных волокон.
Ulva
Ульва многоклеточная с листообразным телом толщиной в две клетки, но длиной до одного метра. Жизненный цикл — это смена поколений. И гаплоидное, и диплоидное поколения выглядят одинаково ( изоморфных ).
Тубулиниды
Рис. 7. Amoeba proteus X 100
Тубулиниды перемещаются с помощью цитоплазматических расширений, называемых псевдоподиями . Они питаются, фагоцитируя (поглощая) свою добычу.Тубулиниды встречаются в почве, морской и пресной воде. Amoeba proteus (рис. 7) водится в пресной воде.
Передвижение амеб и фагоцитоз (видео)
14 Протиста | Лабораторное руководство для SCI103 «Биология I» в муниципальном колледже Роксбери
Протисты — это любые эукариотические организмы, не являющиеся животными, растениями или грибами. Протисты не образуют естественную группу или кладу, но часто для удобства сгруппированы вместе. В популярной схеме пяти королевств, предложенной Робертом Уиттакером в 1969 году, протисты составляют королевство под названием Протиста, состоящее из «организмов, которые являются одноклеточными или одноклеточно-колониальными и не образуют тканей.Некоторые простейшие являются значительными паразитами животных (например, пять видов паразитического рода Plasmodium вызывают малярию у людей, а многие другие вызывают аналогичные заболевания у других позвоночных), растений (оомицет Phytophthora infestans вызывает фитофтороз картофеля) или даже других простейших. Патогены-протисты разделяют многие метаболические пути со своими эукариотическими хозяевами. Это чрезвычайно затрудняет разработку терапевтических мишеней — лекарство, которое наносит вред простейшим паразитам, также может нанести вред его животному / растению-хозяину.
Термин протисты впервые был использован Эрнстом Геккелем в 1866 году. Протисты традиционно подразделялись на несколько групп на основе сходства с «высшими» царствами, например:
Таксономия протистов постоянно меняется. Новые классификации пытаются представить монофилетические группы на основе морфологической (особенно ультраструктурной), биохимической (хемотаксономия) и информации о последовательности ДНК (молекулярные исследования). Однако иногда наблюдаются расхождения между молекулярными и морфологическими исследованиями.
Просмотр живых организмов
Амеба протей
Amoeba proteus (рис. 14.1) — амеба, тесно связанная с гигантскими амебами. Это небольшое простейшее использует щупальца, называемые псевдоподиями, для перемещения и фагоцитоза более мелких одноклеточных организмов (которые могут быть больше по размеру, чем у амеб), которые заключены внутри цитоплазмы клетки в пищевую вакуоль, где они медленно разрушаются ферментами. Он населяет пресноводную среду и питается другими простейшими, водорослями, коловратками и даже другими более мелкими амебами.Из-за фитохромов A. proteus может проявляться под микроскопом в различных цветах (часто желтом, зеленом и пурпурном).
Рисунок 14.1: Amoeba proteus.
Paramecium caudatum
Paramecium caudatum (рис. 14.2) — одноклеточный инфузорийный эукариот. Они могут достигать 0,25 мм в длину и покрыты мельчайшими волосковидными органеллами, называемыми ресничками. Реснички используются для передвижения и питания. P. caudatum питаются бактериями и мелкими эукариотическими клетками, такими как дрожжи и жгутиковые водоросли.В гипотонических условиях (пресная вода) клетка поглощает воду путем осмоса. Он регулирует осмотическое давление с помощью сократительных вакуолей, похожих на мочевой пузырь, собирая внутреннюю воду через свои звездообразные радиальные каналы и вытесняя избыток через плазматическую мембрану. Двигаясь по воде, они движутся по спиральной траектории, вращаясь по длинной оси.
Paramecium имеют два ядра (большое макроядро и одно компактное микроядро). Они не могут выжить без макронуклеуса и не могут воспроизводиться без микроядра.Как и все инфузории, парамеции размножаются бесполым путем, путем бинарного деления. Во время размножения макронуклеус расщепляется по типу амитоза, а микроядра подвергаются митозу. Затем клетка делится поперечно, и каждая новая клетка получает копию микроядра и макронуклеуса.
Деление может происходить как часть нормального вегетативного клеточного цикла. При определенных условиях ему может предшествовать самооплодотворение (автогамия) или может следовать конъюгация, сексуальный феномен, при котором парамеции совместимых типов спаривания временно сливаются и обмениваются генетическим материалом.Во время конъюгации микроядра каждого конъюганта делятся мейозом, и гаплоидные гаметы переходят от одной клетки к другой. Затем гаметы каждого организма сливаются с образованием диплоидных микроядер. Старые макронуклеусы разрушаются, а новые развиваются из новых микроядер. Без омолаживающих эффектов автогамии или конъюгации Paramecium стареет и умирает. Только противоположные типы спаривания или генетически совместимые организмы могут объединяться в конъюгацию.
Рисунок 14.2: Paramecium caudatum.
Эвглена
Euglena (рис. 14.3) — род одноклеточных жгутиковых эукариот. Это самый известный и наиболее широко изученный представитель класса Euglenoidea, разнообразной группы, включающей около 54 родов и не менее 800 видов. Виды эвглены водятся в пресных и соленых водах. Они часто многочисленны в тихих внутренних водах, где они могут цвести в количестве, достаточном, чтобы окрасить поверхность прудов и канав в зеленый (E. viridis) или красный (E. sanguinea).При питании в качестве гетеротрофа эвглена поглощает питательные вещества за счет осмотрофии и может выжить без света на диете, состоящей из органических веществ, таких как экстракт говядины, пептон, ацетат, этанол или углеводы. Когда солнечного света достаточно для питания путем фототрофии, он использует хлоропласты, содержащие пигменты хлорофилл а и хлорофилл b, для производства сахаров путем фотосинтеза. Хлоропласты эвглены окружены тремя мембранами, в то время как у растений и зеленых водорослей (среди которых ранние систематики часто помещали эвглену) только две мембраны.Этот факт был воспринят как морфологическое свидетельство того, что хлоропласты Эвглены произошли от эукариотической зеленой водоросли. Таким образом, интригующее сходство между эвгленой и растениями могло возникнуть не из-за родства, а из-за вторичного эндосимбиоза. Молекулярно-филогенетический анализ подтвердил эту гипотезу, и теперь она является общепринятой.
Рисунок 14.3: Эвглена.
Peranema
Peranema (рис. 14.4) — это род свободноживущих жгутиконосцев, насчитывающий более 20 признанных видов, размер которых варьируется от 8 до 200 микрометров.Они обитают в пресноводных озерах, прудах и канавах и часто встречаются на дне застойных бассейнов, богатых разлагающимся органическим материалом. Хотя они принадлежат к классу Euglenoidea и морфологически похожи на зеленую эвглену, Peranema не имеет хлоропластов и не может питаться путем аутотрофии. Вместо этого они захватывают живую добычу, такую как дрожжи, бактерии и другие жгутиконосцы, поедая их с помощью жесткого устройства для кормления, называемого «стержневым органом». В отличие от зеленых эвгленид, у них отсутствует как глазное пятно (стигма), так и парафлагелларное тело (фоторецептор), которое обычно связано с этой органеллой.Однако, хотя у Peranema нет локализованного фоторецептора, они обладают светочувствительным белком родопсином и реагируют на изменения света характерным «скручивающим» поведением.
Рисунок 14.4: Перанема.
Хламидомонада
Chlamydomonas (рис. 14.5) — это род зеленых водорослей, состоящих из одноклеточных жгутиконосцев, которые встречаются в стоячей воде и на влажной почве, в пресной воде, морской воде и даже в снегу как «снежные водоросли». Хламидомонада используется в качестве модельного организма для молекулярной биологии, особенно для изучения подвижности жгутиков и динамики хлоропластов, биогенеза и генетики.Одной из многих поразительных особенностей хламидомонады является то, что она содержит ионные каналы (каналы родопсинов), которые непосредственно активируются светом. Эти белки используются в оптогенетике.
Рисунок 14.5: Хламидомонада. Обратите внимание на жгутики.
Гимнодиниум
Gymnodinium — род динофлагеллят. Это один из немногих голых динофлагеллят или видов без панциря (целлюлозных пластин). Динофлагелляты (греч. Dinos «кружащийся» и лат. Flagellum «хлыст, бич») представляют собой большую группу жгутиковых эукариот, составляющих тип Dinoflagellata.Большинство из них составляют морской планктон, но они также обычны в пресноводных средах обитания. Их популяции распределены в зависимости от температуры, солености или глубины. Известно, что многие динофлагелляты являются фотосинтетическими, но большая часть из них на самом деле является миксотрофными, сочетающими фотосинтез с поеданием добычи (фаготрофия). По количеству видов динофлагелляты составляют одну из крупнейших групп морских эукариот, хотя эта группа значительно меньше диатомовых. Некоторые виды являются эндосимбионтами морских животных и играют важную роль в биологии коралловых рифов.Другие динофлагелляты — непигментированные хищники других простейших, а некоторые формы паразитируют.
Пандорина
Pandorina (рис. 14.6) — это род зеленых водорослей, состоящих из 8, 16, а иногда и 32 клеток, скрепленных вместе в основании, образуя глобулярную колонию мешочков, окруженную слизью. Ячейки имеют яйцевидную форму или слегка сужены на одном конце, чтобы иметь форму трапеции или груши. Каждая клетка имеет два жгутика с двумя сократительными вакуолями в основании, глазное пятно и большой чашевидный хлоропласт, по крайней мере, с одним пиреноидом.Колонии координируют движение своих жгутиков, чтобы создать перекатывающееся и плавательное движение. Pandorina показывает начало полярности и дифференциации колоний, наблюдаемых у Volvox, поскольку передние клетки имеют более крупные глазные пятна. Бесполое размножение — это одновременное деление всех клеток колонии с образованием автоколоний, которые высвобождаются в результате желатинизации колониальной оболочки. Половое размножение происходит путем деления каждой клетки колонии на 16-32 зоогамет. Зоогаметы показывают признаки гетерогамии, небольшую разницу в размере и подвижности пар, которые сливаются, образуя зиготу с гладкими стенками.
Рисунок 14.6: Пандорина.
Volvox
Volvox (рис. 14.7) — это род пресноводных водорослей, обитающих в прудах и канавах, даже в неглубоких лужах. Он образует сферические колонии, насчитывающие до 50 000 клеток, о которых впервые сообщил Антони ван Левенгук в 1700 году. Вольвокс отошел от одноклеточных предков примерно 200 миллионов лет назад. Каждая зрелая колония вольвокса состоит из тысяч клеток двух дифференцированных типов клеток: многочисленных жгутиковых соматических клеток и меньшего количества половых клеток, лишенных сомы, которые встроены в поверхность полой сферы или ценобия, содержащего внеклеточный матрикс, состоящий из гликопротеины.Взрослые соматические клетки состоят из одного слоя с обращенными наружу жгутиками. Клетки плавают скоординированно, с четкими передним и задним полюсами. У клеток есть передние глазные пятна, которые позволяют колонии плыть навстречу свету. Бесполая колония включает как соматические (вегетативные) клетки, которые не размножаются, так и большие неподвижные гонидии внутри, которые производят новые колонии в результате многократного деления. При половом размножении образуются два типа гамет. Виды Volvox могут быть однодомными и раздельнополыми.Мужские колонии выпускают многочисленные пакеты спермы, в то время как в женских колониях отдельные клетки увеличиваются и становятся оогаметами, или яйцеклетками.
Вольвокс является факультативно половым и может воспроизводиться как половым, так и бесполым путем. Переход от бесполого размножения к половому может быть вызван условиями окружающей среды и производством фермона, вызывающего половую принадлежность. Устойчивые к высыханию диплоидные зиготы получаются после успешного оплодотворения.
Рисунок 14.7: Volvox.
Оэдогониум
Oedogonium (рисунок 14.8) — род нитчатых зеленых водорослей с неразветвленными нитями толщиной в одну клетку. Оэдогониум может свободно плавать, хотя обычно прикрепляется к водным растениям с помощью фиксатора. Он кажется зеленоватым и обитает в спокойной пресной воде. Оэдогониум может размножаться бесполым путем путем фрагментации нитей через некоторые другие типы неподвижных спор, а также через зооспоры, которые имеют множество жгутиков. Они развиваются в клетке зооспорангия, одна зооспора на зооспорангий. После оседания и потери жгутиков зооспора превращается в нить.Оэдогониум также может размножаться половым путем. Его половой жизненный цикл гаплонический, т. Е. Зигота претерпевает мейоз. Антеридии производят и выпускают сперму, а оогонии производят и выпускают яйцеклетку. Затем яйцеклетка и сперматозоид сливаются и образуют диплоидную зиготу (2n). Затем зигота подвергается мейозу с образованием нитчатой зеленой водоросли, которая является гаплоидной (1n).
Рисунок 14.8: Оэдогониум.
Спирогира
Spirogyra (рис. 14.9; общие названия включают водяной шелк, косы русалки и покровный сорняк) — это род нитчатых хлорофитных зеленых водорослей отряда Zygnematales, названный по спиральному или спиральному расположению хлоропластов, который является диагностическим признаком рода. .Он обычно встречается в пресноводных районах, а в мире насчитывается более 400 видов спирогиры. Ширина спирогиры составляет от 10 до 100 мкм, а длина может достигать нескольких сантиметров. Спирогира может размножаться как половым, так и бесполым путем. При вегетативном размножении происходит фрагментация, и спирогира просто подвергается интеркалярному митозу с образованием новых нитей. Половое размножение бывает двух типов:
1. Скалярное сопряжение требует объединения двух разных нитей, выровненных бок о бок либо частично, либо по всей своей длине.Каждая ячейка из противоположно выровненных нитей испускает трубчатые выступы, известные как трубки конъюгации, которые удлиняются и сливаются, образуя проход, называемый каналом конъюгации. Цитоплазма клетки, действующей как самец, проходит через эту трубку и сливается с цитоплазмой самки, а гаметы сливаются, образуя зигоспору.
2. При латеральной конъюгации гаметы образуются в виде единой нити. Две соседние клетки около общей поперечной стенки дают выпуклости, известные как трубки конъюгации, которые в дальнейшем образуют канал конъюгации при контакте.Мужская цитоплазма мигрирует по каналу конъюгации, сливаясь с женской. В остальном процесс происходит как при скалярном сопряжении.
Существенное различие состоит в том, что скалярное конъюгация происходит между двумя филаментами, а латеральное конъюгация происходит между двумя соседними клетками на одном и том же филаменте.
Рисунок 14.9: Спирогира.
Просмотреть подготовленные слайды
Amoeba proteus (рис. 14.10)
Рисунок 14.10: Amoeba proteus.
Paramecium 4 типа простейших (рисунок 14.11)
Рисунок 14.11: Парамеции и другие протисты.
Paramecium caudatum (рис. 14.12)
Рисунок 14.12: Парамеций.
Парамеций в конъюгации (рис. 14.13)
Рисунок 14.13: Парамеций в конъюгации.
Эвглена (рисунок 14.14)
Рисунок 14.14: Эвглена.
Динофлагеллат (рис. 14.15)
Рисунок 14.15: Динофлагелляты.
Ceratium (рисунок 14.16)
Рис 14.16: Ceratium, динофлагеллята.
Перидиний (Рисунок 14.17)
Рисунок 14.17: Перидиний, динофлагеллята.
Фораминиферы
Foraminifera (рис. 14.18; латинское значение «носители дырок»; неофициально называемые «форамы») являются членами филума или класса амебоидных простейших, характеризующихся: потоком гранулированной эктоплазмы для ловли пищи и других целей; и обычно внешняя оболочка (называемая «тестом») из различных форм и материалов. Большинство фораминифер — морские, большинство из которых живут на донных отложениях или внутри них (т.е., являются бентосными), тогда как меньшие разновидности плавают в толще воды на разной глубине (т. е. являются планктонными). Эти оболочки обычно состоят из карбоната кальция (CaCO 3 ) или агглютинированных частиц осадка. Известно более 50 000 видов, как живых (10 000), так и ископаемых (40 000).
Рисунок 14.18: Фораминиферы.
Радиолярия
Радиолярии (рис. 14.19), также называемые радиозоа, представляют собой простейшие диаметром 0,1–0,2 мм, образующие сложные минеральные скелеты, обычно с центральной капсулой, разделяющей клетку на внутреннюю и внешнюю части эндоплазмы и эктоплазмы.Сложный минеральный скелет обычно состоит из кремнезема. Они встречаются в виде зоопланктона по всему океану, а их скелетные остатки составляют значительную часть покрытия дна океана в виде кремнистого ила.
Рисунок 14.19: Радиолярии.
Диатомовые водоросли
Диатомовые водоросли (рис. 14.20) — основная группа микроводорослей и один из наиболее распространенных типов фитопланктона. Диатомовые водоросли являются производителями в пищевой цепочке. Уникальная особенность клеток диатомовых водорослей заключается в том, что они заключены в клеточную стенку из кремнезема (гидратированного диоксида кремния), называемую панцирем.Эти панцири очень разнообразны по форме, но обычно почти двусторонне симметричны, отсюда и название группы. Эти раковины используются людьми как диатомовая земля, также известная как диатомит. Ископаемые остатки предполагают, что они возникли в ранний юрский период или раньше. Только мужские гаметы центрических диатомовых способны перемещаться посредством жгутиков. Сообщества диатомовых водорослей — популярный инструмент для мониторинга состояния окружающей среды, прошлого и настоящего, и обычно используются при исследованиях качества воды.
Рисунок 14.20: Диатомовые водоросли.
Trypanosoma cruzi и Trypanosoma brucei gambiense
Trypanosoma cruzi — вид паразитических эвгленоидов. Среди простейших трипаносомы обычно несут ткань в другом организме и питаются кровью (в первую очередь), а также лимфой. Такое поведение вызывает заболевание или вероятность заболевания, которая зависит от организма: например, трипаносомоз у людей (болезнь Шагаса в Южной Америке).Паразитам нужен организм-хозяин, и триатомин, вызывающий гематофаг (по описанию «жук-убийца», «жучок с конусным носом» и «жучок-целующий»), является основным переносчиком в соответствии с механизмом заражения. Триатомину нравятся гнезда позвоночных животных для укрытия, где он кусает и высасывает кровь для пропитания. Отдельные триатомины, инфицированные простейшими в результате других контактов с животными, передают трипаносомы, когда триатомин откладывает свои фекалии на поверхности кожи хозяина, а затем кусает. Проникновение инфицированных фекалий дополнительно облегчается из-за того, что человек или животное-хозяин царапают место укуса.
Trypanosoma brucei (рис. 14.21) — это вид паразитарных кинетопластид, принадлежащих к роду Trypanosoma. Паразит является причиной трансмиссивного заболевания позвоночных животных, включая человека, переносимого видами мухи цеце в Африке к югу от Сахары. У человека T. brucei вызывает африканский трипаносомоз или сонную болезнь. У животных он вызывает трипаносомоз животных, также называемый наганой у крупного рогатого скота и лошадей. T. brucei традиционно подразделяется на три подвида: T.б. brucei , Т. б. gambiense и T. b. rhodesiense . Первый — паразит позвоночных, не являющихся человеком, а два последних — известные паразиты человека.
Рисунок 14.21: Trypanosoma brucei gambiense среди эритроцитов.
Плазмодий вивакс
Plasmodium vivax (рис. 14.22) — простейший паразит и патоген для человека. Этот паразит является наиболее частой и широко распространенной причиной рецидивирующей (доброкачественной третичной) малярии, P.vivax — один из пяти видов малярийных паразитов, которые обычно инфицируют людей. Хотя он менее вирулентен, чем Plasmodium falciparum, самый смертоносный из пяти малярийных паразитов человека, малярийные инфекции P. vivax могут привести к тяжелым заболеваниям и смерти, часто из-за патологически увеличенной селезенки. P. vivax переносится самкой комара Anopheles, поскольку кусает только самка этого вида.
Рис. 14.22: Мерозоиты и трофозоиты Plasmodium vivax (кольцевая стадия).
Смешанные зеленые водоросли (рис. 14.23)
Рисунок 14.23: Различные зеленые водоросли.
Хламидомонада (рис. 14.24)
Рисунок 14.24: Хламидомонада. Обратите внимание на жгутики.
Пандорина (Рисунок 14.25)
Рисунок 14.25: Пандорина.
Volvox (Рисунок 14.26)
Рисунок 14.26: Volvox.
Половые стадии вольвокса (рис. 14.27)
Рисунок 14.27: Половые стадии вольвокса.
Spirogyra (рисунок 14.28)
Рисунок 14.28: Спирогира.
Зооспоры Oedogonium (рис. 14.29
Рисунок 14.29: Зооспоры Oedogonium.
Oedogonium macrandous (рис. 14.30)
Рисунок 14.30: Оэдогониум.
Фукус мужской и женский концептакл
Fucus — это род бурых водорослей, обитающих в приливных зонах каменистых побережий почти по всему миру. Он имеет относительно простой жизненный цикл и дает только один тип слоевища, который вырастает до максимального размера 2 м.Слоевище многолетнее с неправильной или дискообразной опорой или с гаптерами. Прямостоячая часть слоевища дихотомическая или подпорчинно разветвленная, уплощенная и с отчетливой средней жилкой. Заполненные газом пневмоцисты (воздушные пузырьки) у некоторых видов присутствуют парами, по одной по обе стороны от средней жилки. Гаметангии развиваются в концептаклах, встроенных в приемники на вершинах последних ветвей. Они могут быть однодомными или раздельнополыми.
Плодородные полости, концептуальные клетки, содержащие репродуктивные клетки, погружены в сосуды у концов ветвей.После того, как мейозные оогонии и антеридии продуцируются и высвобождаются, следует оплодотворение, и зигота превращается непосредственно в диплоидное растение. Это можно рассматривать как аналог жизненного цикла цветущего растения, но у водорослей оогонии высвобождаются и удобряются в море, в то время как у цветущих растений семяпочки оплодотворяются, будучи прикрепленными к родительскому растению, а затем высвобождаются в виде семян.
Фукус мужской концептакль (Рисунок 14.31)
Рисунок 14.31: Мужской концептакль фукуса
Женский концептакль Fucus (Рисунок 14.32)
Рисунок 14.32: Женский концептакль Fucus
Полисифония
Полисифония (рис. 14.33)
это род нитчатых красных водорослей, насчитывающий около 19 видов на побережье Британских островов и около 200 видов по всему миру.
Рисунок 14.33: Полисифония.
Стемонит
Stemonitis (рис. 14.34) — особый род слизистой плесени, встречающийся во всем мире (кроме Антарктиды).Для них характерны высокие коричневые спорангии, опирающиеся на тонкие стебли, которые растут гроздьями на гниющей древесине.
Рисунок 14.34: Стемонит.
Сапролегния
Saprolegnia (рис. 14.35) одновременно является сапротрофом и некротрофом. Обычно они питаются отходами рыб или другими мертвыми клетками, но также могут воспользоваться ранеными существами. Инфекция, известная как оомикоз. Сапролегния толерантна к широкому диапазону температур, от 3 ° C до 33 ° C, но чаще встречается при более низких температурах.Хотя он чаще всего встречается в пресной воде, он также переносит солоноватую воду и даже влажную почву. Нити сапролегнии (гифы) длинные с закругленными концами, содержат зооспоры. Сапролегния обычно путешествует колониями, состоящими из одного или нескольких видов. Сначала они образуют массу отдельных гиф. Когда масса гиф становится достаточно большой, чтобы ее можно было увидеть без использования микроскопа, ее можно назвать мицелием.
Рисунок 14.35: Сапролегния.
Он имеет диплоидный жизненный цикл, включающий половое и бесполое размножение.В бесполой фазе спора Saprolegnia выделяет зооспоры. Через несколько минут эта зооспора прорастет, прорастет и выпустит другую зооспору. Эта вторая зооспора имеет более длинный цикл, во время которого происходит наибольшее распространение; он будет продолжать энцистировать и выпускать новую спору в процессе, называемом полипланетизмом, до тех пор, пока не найдет подходящий субстрат. Когда подходящая среда будет найдена, волосы, окружающие спору, будут сцепляться с субстратом, так что может начаться фаза полового размножения. Также именно на этой стадии полипланетизма сапролегнии способны вызывать инфекцию; у наиболее патогенных видов есть крошечные крючки на концах волос, чтобы повысить их инфекционную способность.После прочного прикрепления половое размножение начинается с производства мужских и женских гаметангиумов, антеридий и оогониумов соответственно. Они соединяются и соединяются через трубки для удобрения. Образовавшаяся зигота называется ооспорой.
17.6: Жизненный цикл фукуса — Biology LibreTexts
- Последнее обновление
- Сохранить как PDF
- Авторы и авторства
Наш модельный организм для жизненного цикла Phaeophyta — это Fucus (водоросль), который, как и его родственник Saprolegnia, имеет дипломатический жизненный цикл.
Обратите внимание на отображаемое слоевище Fucus . Обратите внимание на дихотомическое ветвление (разветвление на две равные ветви) и вздутые сердцевидные репродуктивные кончики ветвей. Эти вздутые кончики ветвей называются сосудами . Емкости покрыты небольшими выпуклостями, каждая с порами в центре выпуклости, именуемыми ostiole . Бугорки — это концептакулов , камеры, в которых находятся мужские и женские гаметангии.
Рис. \ (\ PageIndex {1} \): Fucus conceptacle
Просмотрите подготовленные слайды поперечного сечения через приемник Fucus , чтобы рассмотреть концептуальные элементы.Мужские и женские гаметангии размещаются в камере концептакла. Антеридии представляют собой разветвленные структуры, похожие на небольшие деревья. Они продуцируют сперматозоидов, с гетероконтами жгутиков.
Оогонии представляют собой шаровидные структуры, разделенные на секции, в результате чего получается яиц . Яйцеклетки будут оплодотворены спермой, которая проходит через устиол, образуя диплоидную зиготу , которая будет выпущена в морскую воду. Эта зигота вырастет путем митоза в многоклеточный диплоидный слоевище.Обозначьте выделенные жирным шрифтом структуры на схеме жизненного цикла на следующей странице.
В жизненном цикле Fucus ниже пометьте все важные структуры, укажите, где происходят мейоз и оплодотворение, и по-разному раскрасьте гаплоидные и диплоидные ткани. Нарисуйте стрелки, показывающие, где происходит митоз.
Рисунок \ (\ PageIndex {1} \): Диаграмма жизненного цикла Fucus
Авторы и авторство
.